 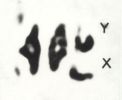    
|


|
|
Goldschmidt, Elizabeth. 1999, Class demonstration of allelic complementation at the ma-1 locus and maternal effect of the ma-1+ gene.
Dros. Inf. Serv. 82: 125-126 View PDF
Moree, Ray. 1999. Simple demonstration of modified ratios using b and e.
Dros. Inf. Serv. 82: 126. View PDF
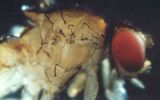
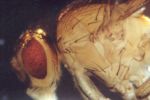
In D. melanogaster intra-locus complementation can be observed at the morphological level in heterozygotes for different pseudo-alleles at any one of a series of loci, such as fa-spl or m-dy. Complementation at the ma-1 locus which has been demonstrated at the enzymatic level (Glassman, 1960) may be preferable for class work. Heterozygous females of the constitution ma-1/ma-1bz synthesize considerable quantities of the enzyme xanthine dehydrogenase (XDH) which is virtually absent in either homozygous mutant. Although this effect is amenable to investigation with advanced enzymological and immunological techniques, its main features may be illustrated by one-dimensional chromatography of mutant flies, following Hadorn and Mitchell (1951).
One of the immediate products of XDH activity is isoxanthopterine (IX). Neat separation of IX from bodies of female flies may be obtained by chromatography of Malphighian tubules. the Malphighian tubules of 8 females are applied to each start point of a sheet of filter paper. After development in (2:1) propanol:5% ammonia solution in a darkened room, the IX spot with its characteristic violet fluorescence will be seen under a U.V. lamp (360 mm), in chromatograms of Malphighian tubules derived from normal females and of ma-1/ma-1bz heterozygotes, but not in those of either homozygous mutant type. Partial complementation of the amount of red pigment (drosopterine), the synthesis of which is also correlated with the XDH level, can be demonstrated by chromatography of single female heads of the same four genotypes.
The maternal effect of ma-1+ females on the XDH content of their mutant offspring may be shown by chromatography of testes of mutant males. The testes of two males applied to each start point yield large spots of IX. The maternally affected males should be obtained from the following crosses:
1) y ma-1+:= X ma-1
2) y ma-1+:= ; ry/ry X ma-1;+/+
The latter cross demonstrates that ry/ry females although devoid of XDH can exert a maternal effect on the production of the enzyme in their ma-1 sons.
For technical details and further references see: Glassman, E., 1960, Science 131: 1810; Glassman, E., and J. McLean 1962, Proc. Nat. Acad. Sci. 48: 1712; Hadorn, E., and H.K. Mitchell 1951, Proc. Nat Acad. Sci. 37: 650.
Laboratory experiments relating to modified ratios and genic interaction can be made both simple and surprising by using stocks of b and e. That the F1 is wild type is surprising to many. The F2 is classified by most students into wild type and “dark”, in 9:7 ratio if the sample size is large enough. But some students detect what they consider different degrees of darkness, so the possibilities of getting a 9:3:4 or a 9:6:1 ratio are pointed out: it can also be indicated that if a simple chemical test were available it might even be possible to recognize a 9:3:3:1 ratio. That the ratio of wild to dark may be about 1:1 in F2 progeny and 1:3 in testcross progeny from the cross F1 X b/b;e/e is usually somewhat surprising, too. The results emphasize the way in which inferences as to interaction, epistasis, etc., depend upon the possibilities of discriminating among the individual progeny of a cross.