
Chao, T.H. , A. Gheta, Y.K. Kim, R. Cella, and L. Ehrman. 2001. Male pheromones of Drosophila paulistorum. Dros. Inf. Serv. 84: 50-54.
|
|
|
|||
|
|
||||
Male pheromones of Drosophila paulistorum.
Chao,
T.H. 1, A. Gheta1, Y.K. Kim2, R. Cella1,
and L. Ehrman1.
1 Division of Natural Sciences, Purchase
College, State University of New York, Purchase, NY, 10577; 2 Department of Genetics, University
of Georgia, Athens, GA 30602.
Abstract
Some
insects, including flies, employ a cuticular pheromonal communication system
to regulate mating behavior. We
studied the cuticular lipids of both males and females of the six Drosophila
paulistorum semispecies. Cuticular
compounds were extracted with hexane, separated on a silica gel column, and
analyzed using GC/MS. All six
semispecies have basically similar lipid patterns, except for two components
which are found only in males. Methyl tetradecenoate
(A) and docosenyl acetate (B) were found in the fraction eluted by 5% ether/hexane,
and are present in A/B ratios varying from 0.22-1.33 for the six semispecies.
Comprehensive bioassays established the pheromonal role of these two
esters in D. paulistorum males.
Introduction
It is known that some compounds, found in Drosophila cuticles, are pheromones that play critical roles in mating preference, showing intra- and inter-specific variation. For the much studied species group D. melanogaster, the major female contact pheromone is (Z, Z)-7,11-heptacosadiene; whereas, the same compound was not found in sexually mature males. Instead, (Z)-vaccenyl acetate, possessing antiaphrodisiac properties (Jallon, 1984), is synthesized in the mature males. Although almost identical chemical composition of cuticular hydrocarbons were found among members of this species, some quantitative variations have been found between geographic populations (Jallon and Pechine,1989).
No
chemical analysis of the cuticular compounds of the D. paulistorum group has been reported, although literature exists
on the biology of their behavior and mating preferences (Ehrman, 1965, 1995).
The six semispecies, including Andean Brazilian (AB), Amazonian
(AM), Centroamerican (CA), Interior (IN), Orinocan (OR), and Transitional
(TR), all native of South America, are mostly allopatric. But in some places their geographic ranges
overlap, and two or more semispecies may coexist sympatrically without interbreeding.
Here we report our findings
from gas chromatography/mass spectrometry (GC/MS) analyses and from bioassays
of cuticular compounds.
Methods
For most studies, 3-day old, sexually mature male and female virgin flies were used. Body weight averages 0.5 mg. In a typical experiment, approximately 200 3-day-old virgin flies per sex were killed by storing at -70°C. The flies were then washed in hexane to obtain their cuticular lipid extract. This hexane extract was evaporated to dryness under nitrogen and then reconstituted in hexane to approximately 10 μl for analyses by GC/MS. Tetracosane was added as the internal standard. Both electron impact (EI) and chemical ionization (CI)/methane methods were used. Analyses were carried out on an HP 5890A/5971A instrument and a 25 m ´ 0.32 mm ´ 0.52 μm AT-1 column, programmed between 150ºC and 300ºC at 5°C/min or a 30 m ´ 0.25 mm DB-1 column, programmed from 50ºC (1min) to 320ºC at 6ºC/min.
Hexane extract was separated into fractions according to polarity on a mini glass column packed with silica gel, eluted with solvents of increasing polarity: hexane, 5% ether/hexane, 10% ether/hexane, and 50% ether/hexane.
Bioassay
of male cuticular extracts involves direct mating observations. A sample of
the hexane extract or column chromatographic fraction obtained from 20 flies
was applied to a strip of filter paper, which was allowed to stand until solvent
had completely evaporated. Twenty
virgin females were then exposed to this test material for 5 min. Subsequently,
the exposed females were placed in a mating chamber (Elens and Wattiaux, 1964)
together with 20 unexposed, clean females (wings clipped to facilitate identification),
plus 40 males. All this was accomplished
without anesthetization. Mating patterns were recorded until all flies copulated.
Results
and Discussion
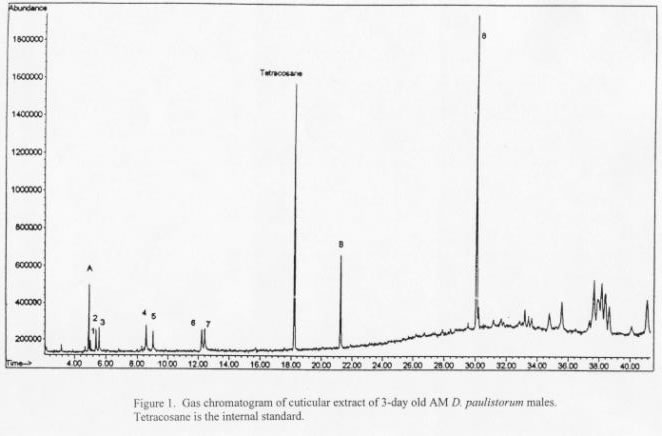
Data from GC/MS showed that the major cuticular lipid components in all six semispecies of both sexes are qualitatively the same, but all males possess the same two components that are completely absent in their female counterparts. Figure 1 is the typical gas chromatogram of the male lipid extracts. Only major peaks are shown, and the peaks that are present in males only are labeled “A” and “B”.
Peaks A and B and Peaks 2-7 are found in chromatographic fractions eluted with 5% ether/hexane and 10%- 50% ether/hexane, respectively. Hexane fraction contained Peaks 1 and 8 and the clusters of peaks after #8. The most abundant peaks are Peak B and #8.
Mass spectra data confirm that Peaks 1 and 8 are hydrocarbons, n-heptadecane and 2-methyl triacotane (C31H64), and that Peaks 2-7, the more polar compounds, are fatty acids ranging from C14 to C18. No serious attempts have been made to identify the clusters of peaks after Peak 8; mass spectra suggest they are long chain hydrocarbons.
Compound A has been identified as methyl tetradecenoate (C15H28O2, MW 240), based on its mass spectrum, which is identical to that of 11-methyl tetradecenoate in the NIST98 MS library. However, the position of the double bond still needs to be confirmed via derivatization experiments.
Figure 2 gives the EI mass spectrum of compound B. No known literature references can be found for this compound. The EI spectrum shows a typical low mass ion series characteristic of an alkene functionality, a prominent peak at m/z 43 (due to CH3C=O+), and a highest mass ion peak at m/z 306. CI mass spectrum gave strong peaks at m/z 367 (MH+) and 307 (MH+ - 60). Mass spectral data therefore suggest compound B is docosenyl acetate (C24H46O2, MW 366). The absence of molecular ion peak in the EI spectrum is consistent with the behavior of an acetate. Again, additional experiments will be necessary in order to determine the position of the double bond as well as to confirm the assignment.
A comprehensive bioassay was carried out to determine whether the two male specific compounds indeed have pheromonal properties. The premise of the experimental design is to determine if the presence of these compounds, on the cuticles of virgin females, significantly reduces male courtship. Number of copulae are recorded for unexposed, clean virgin females vs. those virgin females exposed to test materials including a blank (hexane), whole extracts, and column chromatographic fractions. Forty matings, lasting approximately 30 - 40 minutes, were scored for each experiment. For blank and total extract, observations, reported in Table 1, represent the average of two replicates.
For rotated view of figure 1 below click here.

|
Table 1. Number of D.
paulistorum
matings observed during the first half (20) of all possible 40 matings
by exposed females.
|
Table 1 results confirm that compounds acting as antiaphrodisiacs, the male pheromones, are present in the 5% ether/hexane fraction, and, of course, in the total extract. It is of interest to note that, in both D. paulistorum and D. melanogaster, the major antiaphrodisiacs are esters and not simple hydrocarbons.

Figure
2. EI mass spectrum of compound
B (docosenul acetate).
Figure 3 shows preliminary data on amounts of these two male pheromone compounds as a function of developmental stage. Amounts (in ng) were calculated by comparing the area of the GC peak due to the compound of interest against that of the internal standard, tetracosane. Intervening data points, between 3 and 10 days, are needed, as well as replicate ones. Nonetheless, those compounds identified here as pheromones show the most significant increases at sexual maturity (2-3 days).
|
Figure 3. Amount of cuticular pheromonal compounds
vs. age of AM D. paulistorum
males. --5--
C15H28O2, --= --, C24H46O2 |
We also found that in 3-day old males, relative amounts of A and B in AB, IN, AM, TR, OR, and CA semispecies expressed in the ratio of A/B, arranged in increasing order, are 0.22, 0.25, 0.40, 0.47, 0.63, and 1.33, respectively, which does not seem to correlate with one putative phylogenetic order: TR (ancestor) > IN> CA > OR > AB > AM., proposed by Gleason et al. (1998).
Conclusions
Two male-specific compounds, methyl tetra-decenoate (C15H28O2) and docosenyl acetate (C24H46O2), have been identified in all six semi-species of D. paulistorum, though with significant quantitative differences. These esters exhibit antiaprodisiac properties. We hope, as we gather more comprehensive chemical information about these semispecies, we will be able to identify those genetic factors that might play an important role in the “speciation of D. paulistorum pheromonally.”
Acknowledgments: We wish to thank Jerome Klun of USDA for samples of methyl 11-tetradecenoate. Chemical Ionization mass spectra were obtained at the Mass Spectrometry Facility at University of California, Riverside. This research was funded by a Merck/AAAS USRP award and a grant from Texaco, Inc.
References: Ehrman, L., 1965, Evolution 19: 459-464; Ehrman, L., I. Perelle, and J. Factor 1995, Genetics of Natural Populations, The Continuing Influence of Theodosius Dobzhansky (L. Levine, ed.), Chapter 18: 241-261. Columbia University Press: New York; Elens, A.A., and J.M. Wattiaux1964, Dros. Inf. Serv. 39: 118-119; Gleason, J.M, E.C. Griffith, J.R. Powell 1998, Evolution 52(4): 1093-1103; Jallon, J.-M., 1984, Behaviour Genetics 14: 441-476; Jallon, J.-M., and J.M. Pechine 1989, C.R. Acad. Sci. Paris 309: 1551-1556.

{kind=link}