
Valente, V.L.S., C. Rohde, V.H. Valiati, N.B. Morales, and B. Goñi. 2001. Chromosome inversions occurring in Uruguayan populations of Drosophila willistoni.. Dros. Inf. Serv. 84: 55-59.
|
|
|
|||
|
|
||||
Chromosome inversions occurring in Uruguayan populations of Drosophila willistoni.
Valente1,
V.L.S., C. Rohde1, V.H. Valiati1,2, N.B. Morales1,
and B. Goñi3. 1Departamento de Genética,
Instituto de Biociências, Universidade Federal do Rio Grande do Sul.
Caixa Postal 15053. CEP 91501-970. Porto Alegre, RS, Brazil. E-mail: vera.gaiesky@ufrgs.br.
2 Laboratório de Biologia Molecular, Centro de Ciências
da Saúde, Universidade do Vale do Rio dos Sinos. Caixa Postal 275.
CEP 93022-000 São Leopoldo, RS, Brazil. 3Sección
Genética Evolutiva, Instituto de Biologia, Facultad de Ciencias, Universidad
de la Republica, Iguá 4225, Montevideo 11400, Uruguay. E-mail:bgoni@fcien.edu.uy.
Reports of Drosophila willistoni
in several localities of Uruguay (Goñi et al., 1997,
1998), near to the southern limit of the geographical distribution of the
species (Spassky et al., 1971)
moved us to investigate the putative paucity of gene arrangements reported
for this species in their borders (Townsend, 1952;
Da Cunha et al., 1950,
1959), as well as the occurrence
of novel or unique gene arrangements.
Only a register exists of the chromosome inversion polymorphism of
D. willistoni in this Country,
from the locality of Melo city,
Northeast Uruguay, reported by Da Cunha and Dobzhansky (1954). The present report aims to start the characterization of the
chromosome inversion polymorphism of D. willistoni populations from Uruguay. Samples of natural populations of D. willistoni were obatined at nine collecting sites (three sites
in the Capital Montevideo: 34o 48’S; 56o 11’W)
and one in each of the following localities: Santa Lucia del Este, Canelones (34o
50’S; 56o 15’W);
Cerro El Toro, Maldonado (34o 45’ S; 55o 14’W); Boca del Sarandi, Rocha (33o 58’S; 53o
43’W); Sauce de Cebollatí,
Lavalleja (33o45’S; 54o33’W); Tacuarembó,
Tacuarembó (31o 41’S; 55o 59’W);
and Arroyo Gajo de Lunarejo, Rivera (31o 06’S; 56o
00’W), in Uruguay. The
flies were obtained as adults around
conventional banana baits and/or as adults emerging from rotten fruits of
Syagrus romazoffiana, Gingko biloba, Doryalis caffra, Butia capitata,
and Citrus sinensis. Third
instar larvae of isofemale lines of each sample were dissected and processed
according to Ashburner (1967).
All samples were chromosomally
polymorphic for paracentric inversions. In contrast to the situation that occurs in D. willistoni
populations of the neighbouring Brazilian Southern State Rio Grande do Sul
(Valente and Morales, 1985; Valente et al., 1993), both arms of the X chromosome are polymorphic.
In this chromosome, we observed 4 inversions, being two in the left
arm (XL) (Figure 1a,c) and two in the right chromosomal arm (XR) (Figure 1b,d). In the autosomal IIL (the left arm of the second chromosome),
a complex arrangement (D+E), also common in Brazil, plus 5 single inversions
were detected (Figure 2). Four
single heterozygous inversions were detected in the right arm of the
second chromosome (IIR) (Figure 3)Uruguayan populations of D. willistoni,
and six were found in the third acrocentric chromosome
(III) (Figure 4). It is interesting
to note the inversion III N, that seems to be endemic from Uruguay, was only
reported previously in a single larva in the sample from Melo city by Da Cunha
and Dobzhansky (1954). The XR
B1 inversion also appears to be endemic from Uruguay, and there are no previous
registers of its occurrence in other natural populations. The first results suggest that in the
Uruguayan populations, several inversions still segregate, despite the fact
that they need to face an environment very different from those found in the putative
center of the species dispersion
- Central Brazil (Da Cunha et
al., 1950). The uniqueness of certain arrangements and of the Uruguayan
populations of D. willistoni
deserves to be further studied.
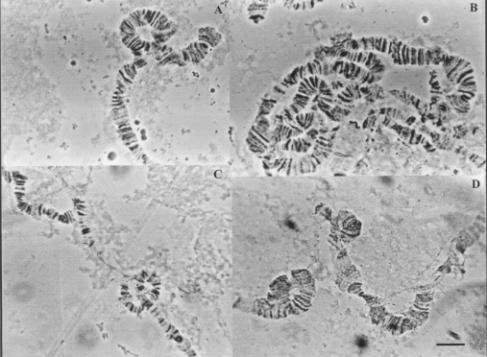
Figure 1. Heterozygous inversions of the X chromosome
of D. willistoni populations from Uruguay.
(a) XL POA 3; (b) XR E; (c) XL A; (d)
XR B1. Bar = 10 mm.
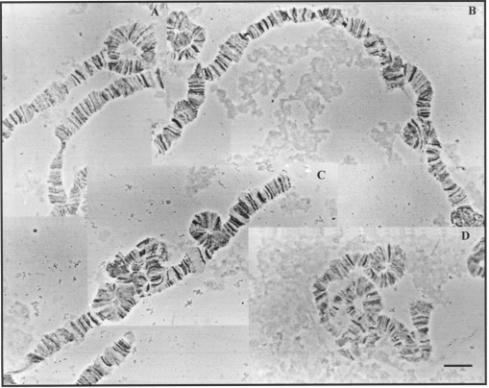
Figure 2. Heterozygous inversions of chromosome
arm IIL of D. willistoni populations
from Uruguay. (a) IIL H; (b) IIL I and IIL B; (c) IIL F and IIL D+E; (d) IIL H, IIL D+E and IIL A. Bar =10 µm.
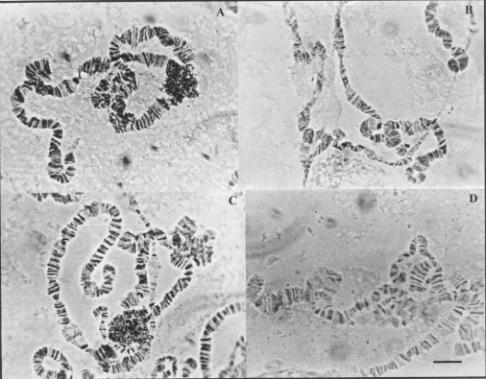
Figure 3. Heterozygous inversions of the chromosome arm IIR of D. willistoni populations from Uruguay. (a) IIR B; (b) IIR C; (c) IIR I; (d) IIR G. Bar = 10 mm.

Figure 4. Heterozygous inversions of the chromosome
III of D. willistoni populations from
Uruguay. (a) III J and III B;
(b) III J; (c) III J and III C; (d) III N; (e) III J+V1; (f)
III M. Bar =10 mm.
Acknowledgments: CSIC (Uruguay); CNPq, FAPERGS, PROPESQ-UFRGS (Brazil).
References: Ashburner, M., 1967, Chromosoma 27: 47-63; Da Cunha, A.B., H. Burla, and Th. Dobzhansky 1950, Evolution 4: 212-235; Da Cunha, A.B., and Th. Dobzhansky 1954, Evolution 8: 119-134; Da Cunha, A.B., Th. Dobzhansky, O. Pavlovsky, and B. Spassky 1959, Evolution 13: 389-404; Goñi, B., M.E. Martínez, and P. Daguer 1997, Revta. Bras. Ent. 41: 89-93; Goñi, B., M.E. Martínez, V.L.S. Valente, and C.R. Vilela 1998, Revta. Bras. Ent. 42: 131-140; Spassky, B., R.C. Richmond, S. Pérez-Salas, O. Pavlovsky, C.A. Mourão, A.S. Hunter, H. Hoenigsberg, Th. Dobzhansky, and F.J. Ayala 1971, Evolution 25: 129-143; Townsend, J.I., 1952, Evolution 6: 428-442; Valente, V.L.S., and N.B. Morales 1985, Revta. Bras. Genet. 8: 167-173; Valente, V.L.S., A. Ruszczyk, and R.A. dos Santos 1993, Revta. Bras. Genet. 16: 307-319.