
Beltrami, M. and R. Godoy-Herrera. 2001. Larval pupation behavior of the mesophragmatica species group of Drosophila. Dros. Inf. Serv. 84: 101-103.
|
|
|
|||
|
|
||||
Larval pupation behavior of the mesophragmatica species group of Drosophila.
Beltrami, M.,1,2 and R. Godoy-Herrera1. 1Programa de Genética Humana, Instituto de Ciencias Biomédicas, Facultad de Medicina, Universidad de Chile; 2Departamento de Biología, Facultad de Ciencias Básicas, Universidad Metropolitana de Ciencias de la Educación.
The mesophragmatica species group of Drosophila includes at least six sibling species (Brncic and Koref-Santibañez, 1957). Their phylogenetic relationships were established by Brncic et al. (1971) and Nair et al. (1971) by chromosomal and isoenzymatic studies. Del Pino and Godoy-Herrera (1999) found that the developmental patterns of larval foraging behavior in the group follow those phylogenetic relationships. Thus, in the mesophragmatica group, changes in ecologically relevant behaviors exhibited by the larvae during the feeding period paralleled genetic changes that occurred in evolutionary history of the group. Less known is whether changes in larval dispersal patterns involved in selection of pupation sites follow the established phylogenetic relationships among the species of the mesophragmatica group. This type of investigation may be of importance to understanding the evolution of behavioral patterns related with dispersal in heterogeneous enviroments, and it may reveal how closely related species have solved the problem of gaining access to and using resources like food and space.
The stocks employed in this work originated from flies collected by Brncic (1987): i) D. gasici (Cochabamba, Bolivia), ii) D. brncici (Bogotá, Colombia), iii) D. viracochi (Bogotá, Colombia), iv) D. mesophragmatica (La paz, Bolivia), v) D. gaucha (Buenos Aires, Argentina), and vi) D. pavani (La Florida, Santiago de Chile). They are kept by mass culture in the Faculty of Medicine, University of Chile. The flies were all reared in a constant enviroment at 18°C in half-pint bottles. In this work we used 100 eggs of each of the species above indicated which were distributed on separate cups of 2.0 ´ 2.5 cm filled with nutritive medium. The cups were deposited into transparent perspex boxes (see Godoy-Herrera et al., 1989, for details). The bottoms of the boxes were filled with agar to a depth of 2 cm; then an area of agar of 9.0 ´ 4.5 cm was removed along one side to leave a dry surface. This was filled with yellow sand to a depth of 2 cm. Thus, the rearing cup was surrounded by agar and sand. The mature larvae of each of the species could choose to pupate either on agar, on the sand, or in the cup. The sand was humidified with 5 cc of destilled water. The number of pupae detected outside (the agar and sand) and inside the rearing cup was recorded prior to eclosion. We also recorded the number of pupae from each of the species found on and underneath agar and humid sand. Each experiment was replicated four times. The significance of the differences between number of pupae of D. gasici, D. brncici, D. viracochi, D. mesophragmatica, D. gaucha, and D. pavani found inside and outside the food cup was tested by using the G-test of Independence (Sokal and Rohlf, 1995). To test the significance of differences in the number of pupae found on and underneath the agar and humid sand, we applied a Chi-Square of independence R ´ C test. Before this we carried out unplanned tests of homogeneity for replicates made for each group in each experiment (the R ´ C test of independence, Sokal and Rohlf, 1995).
The statistical analysis showed that there were no significant differences
within a species and among replicates in each experiment, and so we pooled
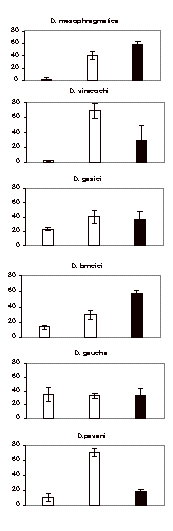
the data as shown below. In Figure
1 is shown the preferences of D. mesophragmatica,
D. viracochi, D. gasici, D. brncici, D. gaucha, and D.
pavani larvae to pupate on agar, moist sand, and the cup.
D. mesophragmatica and D. brncici larvae prefer the sand, while those of D. viracochi and D. pavani pupate on agar. In contrast, D. gasici and D. gaucha larvae seem not to show preferences for the substrates to pupate, as
shown by the percentages of pupae on the cup, agar and sand (Figure 1). The distributional pattern of pupae of
the species is each statistically different with respect to the other ones
(the G-test of independence). The
Chi-Square values all exceed the critical value 5.99, df = 2, p < 0.05.
 |
 |
 |
 |
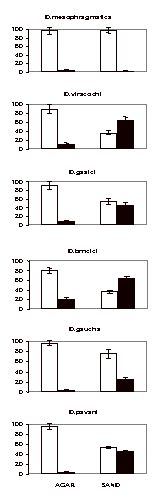
Larval dispersal patterns of Drosophila species previously observed to form puparia could be a part of adaptations of these preadults to distribute in heterogeneous environments. The results of this investigation suggest that in the same enviroments mature larvae of the mesophragmatica species group of Drosophila exhibit different dispersal patterns when searching for pupation sites. The larvae seem to differ in their responses to water content, compactness and texture of the substrates. Physical characteristics of the substrates also affect larval digging behavior, as shown by the proportion of pupae found underneath the agar and sand (Figure 2). Those characteristics could differ among the environments used by each of the species’ lives in wild. Under these conditions, different selective pressures in combination with genetic drift could lead to divergence in larval pupation behavior between the species of the mesophragmatica group.
Acknowledgments: Grant for Doctorate studies FID-Universidad Metropolitana de Ciencias de la Educ.; Postgraduate studies school, Facultad de Medicina Universidad de Chile.
References: Brncic, D., and S. Koref-Santibañez 1957, Evolution 11: 300-310;
Brncic, D., S.N. Puliyampetta, and M.R. Wheeler 1971, University of Texas Publication. Studies
in Genetics 6, 7103, 1-16; Del
Pino, F., and R. Godoy-Herrera 1999, Behaviour 136: 391-409; Godoy-Herrera, R., L. Cifuentes, M. Diaz De Arcaya, M. Fernandez,
M. Fuentes, I. Reyes, and C. Valderrama
1989, Anim. Behav. 37: 820-829; Nair, P., D. Brncic, and K-I. Kojima 1971, University of Texas
Publication. Studies in Genetics 6, 7103, 17-28; Sokal, R.R., and F.J. Rohlf 1995, Biometry. W.H. Freeman and Co, San Francisco.