
Waldrip,W.R., N.T. Takaesu, and S.J. Newfeld. 2001. Identification of blue balancers and mutant collections compatible with hobo element transgenes. Dros. Inf. Serv. 84: 169-172.
|
|
|
|||
|
|
||||
Identification of blue balancers and mutant collections compatible with hobo element transgenes.
Waldrip1,W.R., N.T. Takaesu1, and S.J. Newfeld1,2. 1Department of Biology and 2Graduate Program in Molecular and Cellular Biology, Arizona State University, Tempe, AZ 85287-1501. Corresponding author: Stuart J. Newfeld, Department of Biology, Arizona State University, Tempe, AZ 85287-1501; phone: 480-965-6042; FAX: 480-965-2519; email: newfeld@asu.edu
Introduction
The establishment
of transposable elements as genetic tools has had an enormous impact on our
understanding of organismal biology. Our laboratory has contributed to the development of a genetic
system based upon the hobo element
in Drosophila melanogaster. Relying upon the well-characterized molecular genetics
of the decapentaplegic (dpp) locus, we recently reported that hobo, like the better known P element, is capable of local transposition (Newfeld
and Takaesu 1999). Two strains generated in this study are new alleles of
dpp. The other interest of our laboratory is intercellular signaling
during embryonic development. We wanted to exploit these new dpp alleles to learn more about this complex signaling
molecule. However, a major difference between hobo and P
elements is that many laboratory strains contain endogenous hobo elements. Before we could use these new alleles in
developmental genetics studies of dpp we needed to identify hobo-free
strains of various types (e.g.,
Dpp pathway mutants, blue balancers, UAS and Gal4 strains). Here we report
the results of a survey of 78 strains for the presence of endogenous hobo elements. The survey identified a number of useful
strains. This information is important to the growing number of investigators
interested in utilizing the hobo
system, as an alternative to P,
to address developmental questions.
Methods
Standard methods
of DNA isolation, restriction enzyme digestion, Southern blot preparation
and hybridization were utilized (Newfeld and Takaesu 1999). To identify endogenous
hobo elements, genomic DNA from each strain was digested
with XhoI and hybridized with
a wild type hobo element probe
(pRG2.6X; Blackman et al., 1987).
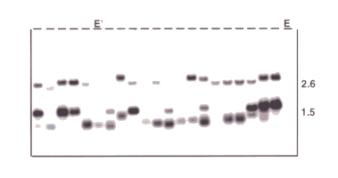
On the autoradiographs, 2.6kb hybridizing fragments represent full-length
(autonomously transposable) hobo
elements. Shorter fragments, typically 1.5kb, represent internally deleted
(non-autonomously transposable) hobo
elements (Blackman et al., 1987).
Strains containing any full-length hobo elements are designated as H strains. Strains containing only internally
deleted hobo elements are designated
as E’ strains. Strains devoid of any hobo elements are designated as E strains.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Figure 1 shows a representative autoradiograph of a genomic Southern blot hybridized with a hobo probe. Of the twenty-two strains shown, twenty are H strains, one is an E’ strain and one is an E strain. Other strains that appear to be E’ strains have faint 2.6kb hybridizing fragments visible on the actual autoradiograph. Table 1 shows the results of our survey of 78 strains. In the survey we found that 58 are H strains (74%), 18 are E strains (23%) and 2 are E’ strains (3%). Despite the high frequency of H strains we were able to identify a number of useful strains including blue balancer strains. In addition, we identified two large mutant collections as possible sources of hobo-free strains for developmental genetic analyses.
|
|
We identified an E’ strain carrying a second chromosome blue balancer - CyO P[ry+; wg.bgal]. This chromosome expresses b-galactosidase in the wingless (wg) pattern of 14 segment polarity stripes and the transgene insertion creates a wg null allele (Kassis et al., 1992). We identified an E’ strain carrying a third chromosome blue balancer - TM3 Sb P[w+; 6.8Xba. bgal]. This chromosome expresses b-galactosidase under the control of a Sex combs reduced hindgut enhancer (Gindhart et al., 1995). Expression from both blue balancers begins at cellular blastoderm and continues throughout development in easily identifiable patterns. We have used these balancers with our new hobo induced dpp alleles for several years and not detected any transposition events (e.g., reversion to dpp+).
In our survey, we tested three mutant strains (cn l(2)IA109 bw sp/ CyO or dJun1, b scws12 pr bw sp/ CyO and ru h th st cu sr e5 kay1 ca/ TM3 Sb Ser) derived from the Nusslein-Volhard and Weischaus (1980) screen for embryonic lethal mutations. Each was designated an E strain suggesting that this mutant collection is a valuable source of hobo-free strains. The only strain (y1 w67c23; mwh red e Med1/ TM3 Sb Ser or l(3)SG70) from the Shearn and Garen (1974) screen for larval/pupal lethal mutations in our survey was designated as an E strain. This suggests, albeit with less confidence, that this mutant collection may also be a source of hobo-free strains. Thus, a large number of embryonic and larval/pupal mutants suitable for use with hobo transgenes may be available for developmental genetics studies.
In summary, we have identified a number of strains that significantly expand the scope of developmental genetic analyses that can be conducted using hobo-mediated transgenes. Investigators interested in further information or in obtaining these strains may contact us.
Acknowledgments: We thank Ann Bradley for assistance in maintaining our strains. This study was supported by a Basil O’Connor Starter Scholar Research Award from the March of Dimes to S. J. N.
References: Blackman, R., R. Grimaila, M. Koehler, and W. Gelbart 1987, Cell 49: 497-505; Gindhart, J., A. King, and T. Kaufman 1995, Genetics 139: 781-795; Kassis, J., E. Noll, E. VanSickle, W. Odenwald, and N. Perrimon 1992, PNAS 89: 1919-1923; Newfeld, S., and N. Takaesu 1999, Genetics 151: 177-187; Nusslein-Volhard, C., and E. Weischaus 1980, Nature 287: 795-801; Shearn, A., and A. Garen 1974, PNAS 71: 1393-1397.
 Figure
1.
Figure
1.