Buratovich, M.A., and S.D. Armer. 2002. The hinge phenotype of In(2L)wgP in Drosophila melanogaster. Dros. Inf. Serv. 85:68-72.
|
|
|
|||
|
|
||||
The hinge phenotype of In(2L)wgP in Drosophila melanogaster.
Buratovich, M.A., and S.D. Armer. Department of Biochemistry, Spring Arbor University, 106 E. Main Street, Spring Arbor, MI, 49283.
Despite the extensive studies of the Drosophila wing and notum, the developmental biology of the wing hinge remains largely uncharacterized. The wingless (wg) gene plays an important role in the development of several structures in the proximal and distal dorsal and ventral hinges (Neumann and Cohen, 1996; Buratovich and Wilder, 2001) . One particular wg allele, In(2L)wgP, shows a dominant outstretched wing phenotype for reasons that remain undetermined. We have examined wing hinges from In(2L)wgP animals to determine if any detectable hinge abnormality exists and to correlate such findings with the mechanism of wg activity in the hinge.
The mutation that originally defined the wg gene was a viable allele that eliminated the wing blade and converted the hinge region into notum (Sharma and Chopra, 1976) . Later mutagenesis screens isolated embryonic-lethal wg alleles that showed a distinctive segment polarity phenotype and other alleles that caused pupal lethality while affecting the development of all imaginal derivatives (Nusslein-Volhard et al., 1984; Baker, 1988a, b) . Several of these x-ray-induced alleles of wg contain severe lesions in the wg-coding region (van den Heuvel et al., 1993) , but one allele in particular, In(2L)wgP, is a regulatory mutation that results from an inversion with breakpoints at 28A1-3 and 32E-F (Baker, 1988b) . The distal breakpoint of In(2L)wgP (hereafter referred to as wgP) at 28A1-3 maps 9-11 kilobases downstream the transcription termination site of the wg gene, presumably within a transcriptional enhancer that controls the imaginal-specific expression of wg (van den Heuvel et al., 1993) . Indeed the designation “wgP” comes from the pupal lethality of this allele in combination with null alleles. Strangely, wgP homozygotes show embryonic lethality and express wg transcripts in seven stripes rather the usual 14 stripes, but wgP heterozygotes also show a dominant adult phenotype that consists of outstretched wings (Baker, 1988b) . Because mutations that affect the structure of the wing hinge often cause flies to hold their wings in the outstretched position at rest (Neumann and Cohen, 1996; Russell, 2000; Buratovich and Wilder, 2001) , an investigation of the wing hinge of wgP heterozygotes might reveal a hinge defect that elucidates the role of Wg in the development of the wing hinge.
Light microscope preparations of hinge hinges were made as previously reported (Buratovich and Wilder, 2001) . In these preparations we used wgP / CyO animals and flies of the same genotype that were outcrossed to Canton S to generate wgP / + heterozygotes. We used these flies with wgP in combination with different chromosomes to eliminate any potential variations in hinge structure due to differing genetic backgrounds. Both stocks showed the outstretched wing phenotype to a great extent. Our wing hinge preparations showed no detectable defects in the dorsal wing hinge (n = 27, data not shown). However, the same preparations of the ventral hinge showed a consistent absence of a structure called the yellow club (YC; data not shown).
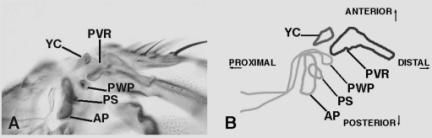
The YC is one of the most conspicuous structures in the ventral wing hinge (Bryant, 1978) . It extends from the cuticle in an anterior direction and lies just proximal to the proximal ventral radius (PVR). The PVR lies at the confluence of the first three wing veins and this thickened flap of wing cuticle points posteriorly. Below the PVR is the pleural wing process (PWP). Extending from the cuticle, toward the PVR, the PWP probably acts with the PVR as a doorstop to prevent the wing from hyperextending in the ventral direction (Dickinson and Tu, 1997) . The PWP extends around an extra pouch of tissue called the axillary pouch (AP), and adjacent to the AP is another sclerite called the pleural sclerite (PS), which is often obscured from view by the AP (Figure 1A-B).
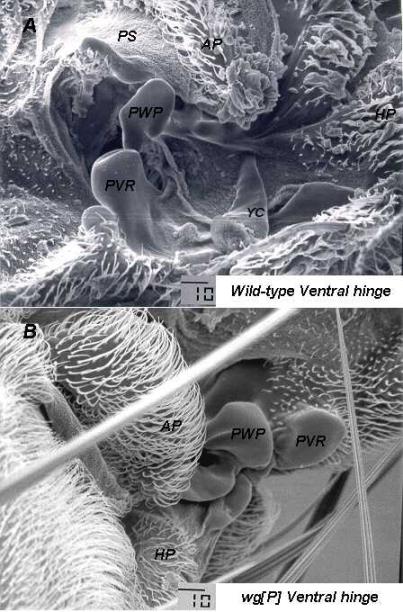
Because of the lack of resolution of light microscopy, we decided to check our light microscopy results with the scanning electron microscope (SEM). We outcrossed wgP / In(2LR)Gla, wgGla-1 animals to Oregon R to generate wgP / + heterozygotes and fixed these adult animals for SEM, according to the procedure of Idle (1970) . Observation of wing hinges from wgP / + heterozygotes (n = 13) with a JEOL JSM T300 scanning electron microscope revealed no detectable defects in the dorsal hinges of these animals (data not shown), but the ventral hinges showed a variety of abnormalities. Some ventral hinges showed deformation of the proximal ventral radius (PVR) and pleural wing process (PWP), even though these structures were always present (data not shown). The YC was consistently missing in these hinges (Figure 2A-B), but in one hinge, the YC was quite small (data not shown). Such diminution could certainly render the YC undetectable during light microscopy.
|
Figure 1. (A) Ventral hinge from a wild-type adult.
Ventrally, the confluence of wing veins 1-3 terminates in the Proximal
Ventral Radius (PVR). Just
proximal to the PVR is the Yellow Club (YC), which is the most conspicuous
ventral hinge structure. Posterior to the YC is a series of plates
and a pouch. The Axillary
Pouch (AP) is an out pocket of the ventral hinge cuticle. Just anterior
to the AP lies the Pleural Wing Process (PWP).
The PWP arches around the AP to form the pleural suture, which
separates the pleura into the mesopleura and the pteropleura. The head of the PWP articulates with
the ventral side of Axillary Sclerite 2 on the dorsal hinge. Posterior to the PWP is the Pleural
Sclerite (PS). The PS is hidden from view in some preparations by
the AP, which overlies and obscures it (Bryant, 1978) . (B) A cartoon
tracing of Figure 1A. |
In situ hybridization of third-instar wing imaginal discs from wgP heterozygotes with wg probes revealed no detectable differences in wg expression when compared to wild-type discs (data not shown). The absence of the YC might have to do with the distal breakpoint of the inversion and nothing to do with the wg gene. A molecular analysis of wgP revertants that no longer show the outstretched wing phenotype should provide some insight into this question.
|
Figure 2. (A) SEM photomicrograph of a ventral hinge from a wild-type Oregon R adult Drosophila. The abbreviations used to label the hinge structures are the same as those used in Figure 1, with the exception of the Humeral Plate (HP), which is a marginal, anterior cuticular plate that spans the anterior margin of the ventral and dorsal wing hinges. The HP lies just distal to the tegula (not shown). (B) SEM photomicrograph of a ventral hinge from a wgP / + adult Drosophila. The YC is missing and the PWP and the PVR are flatter and expanded in appearance.
|
In conclusion the wgP allele of wg causes a dominant wings outstretched adult phenotype that is highly correlated with the elimination or diminution of the Yellow Club in the ventral wing hinge. The significance of this with respect to the role of wg in hinge development or the functional integrity of the wing hinge is not understood at this time.
Acknowledgments: We would like to thank Dow Chemical Corporation
in Central, MI for their kind gift of the JEOL SEM. We would also like to acknowledge the
expertise and contribution of Dr. Fredrick Trexler, professor of physics at
SAU for his help operating and troubleshooting the instrument.