Cook, Kevin R., and Adelaide T.C. Carpenter. 2002. Characterization of Tp(3;3)ME61 and genetic observations on chromosomal regions 96 and 97 of Drosophila melanogaster. Dros. Inf. Serv. 85: 34-37.
|
|
|
|||
|
|
||||
Characterization of Tp(3;3)ME61 and genetic observations on chromosomal regions 96 and 97 of Drosophila melanogaster.
Cook, Kevin R.,1 and Adelaide T.C. Carpenter2. 1Bloomington Drosophila Stock Center, Department of Biology, Indiana University, Bloomington, IN 47405, kcook@bio.indiana.edu; 2Department of Genetics, Cambridge University, Cambridge CB2 3EH, UK, atc12@mole.bio.cam.ac.uk.
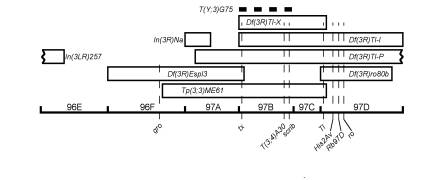
Tp(3;3)ME61 was induced by X-rays on a Df(3L)Pc-101, mwh1 Eip78C101 red1 e4 chromosome in a screen for genetic modifiers of Eip78C. Although the Tp(3;3)ME61 chromosome does not interact with Eip78C, it was retained because it was thought to be a simple deficiency of 96F to 97D. Subsequently, it was found to complement Df(3R)Tl-X (97B;97D1-2), and further cytological analyses showed that the deleted chromosomal segment had been inserted into centric heterochromatin (probably 3Lh). The transposed segment often showed reverse self-synapsis, suggesting it contains an ancestral tandem duplication centered in 97B. The orientation of the transposed segment could not be determined. By cytological criteria, it was impossible to tell whether the deleted chromosomal segment was 96F9-10;97C5-D1 or 96F11-12;97D2-3. The analysis was complicated by the apparent ability of 96F8,9 and 97D1,2 to synapse, which suggests homology between these bands. The complementation tests detailed below indicate that the deletion breakpoints are most likely 96F11-12;97D2-3.
The Df(3R)ME61 component was separated from the Dp(3;3)ME61 component by a H-Pr crossover in Tp(3;3)ME61/H1 Pr1 Bsb1 females. The complementation tests shown in Table 1 were undertaken to define the endpoints of the aberration genetically. The results are depicted graphically in Figure 1. The cytological breakpoints of aberrations used in this study are listed in Table 2.
Cytologically, the proximal euchromatic breakpoint of Df(3R)ME61 lies between the In(3LR)257 breakpoint at 96E2-4 and the proximal breakpoint of Df(3R)Tl-P at 97A2-7 and within Df(3R)Espl3. The gro locus has been mapped to 96F11-14 by in situ hybridization (Hartley et al., 1988), and the complementation of Df(3L)ME61 and gro1 and groC105 suggests that 96F11-12 is more accurate as a proximal breakpoint for Df(3L)ME61 than 96F9-10. While the noncomplementation of Df(3R)ME61 and the deficient inversion In(3R)Na might have allowed the 96F breakpoint of Tp(3;3)ME61 to be localized more precisely, In(3R)Na/Df(3R)Espl3 flies were viable. In fact, the absence of bands in proximal 97A was verified cytologically in polytene preparations from In(3R)Na/Df(3R)Espl3 larvae. This indicates that either the deletion of proximal 97A has no overt phenotypic effect and In(3R)Na and Df(3R)ME61 share a cryptic lethal, or the Df(3R)Espl3 stock carries a transposition of 97A.
|
Figure 1. Tp(3;3)ME61 and the surrounding chromosomal region. Our genetic results strongly suggest
that Tp(3;3)ME61 is
a euploid aberration, so we have used the Tp(3;3)ME61 box to represent both the intact Tp(3;3)ME61 chromosome and the separated Df(3R)ME61 component of the transposition. The thick dashed line indicates the
approximate position of the T(Y;3)G75 breakpoint. |
The distal euchromatic breakpoint of Tp(3;3)ME61 lies between Tl and His2Av, because Df(3R)ME61 failed to complement Tl loss-of-function alleles, but complemented His2Av05146. Tl was mapped to 97D1-2 by in situ hybridization (Ranz et al., 2001) and from the breakpoints of revertants of Tl dominant alleles (Tearle and Nusslein-Volhard, 1987). His2Av was mapped to 97D3-6 by in situ hybridization (Spradling et al., 1999). This suggests that 97D2-3 is the more accurate cytology for the distal Tp(3;3)ME61 breakpoint.
Table 1. Complementation data.
Complementation (C) or noncomplementation
(NC) for visible (v), male sterile (m) or female sterile (f) phenotypes;
|
Table 2. Aberrations
used in this study.
|
The transposed chromosomal segment includes an intact Tl locus, suggesting that it is not grossly deleted at its distal end. Tp(3;3)ME61 complemented Df(3R)Tl-X, placing the breakpoint of the transposition distal to the breakpoint of Df(3R)Tl-X within the Tl-His2Av interval. Likewise, the Tl locus must be intact, because females bearing either Tlr3 or Tlr4 and Tp(3;3)ME61 produced abundant adult progeny. Nevertheless, they produced approximately 10% dead brown-colored embryos, suggesting that the Tl locus is subject to position effect variegation.
The current genome annotation predicts the existence of six genes between Tl and His2Av (Adams et al., 2000). Tp(3;3)ME61/Df(3R)ro80b larvae died attempting to pupariate. Pier Paolo d’Avino, Cambridge University, examined these pseudopupae and reported they had nonexistent or rudimentary imaginal disks and small salivary glands (personal communication). These phenotypes are consistent with disruption, deletion or position effect suppression of genes in the Tl-His2Av interval. Of particular interest are the putative insulin-like growth factor receptors CG6396 and CG6390, and the potential mitotic serine/threonine protein kinase BcDNA:LD09009. Our complementation results between Df(3R)Tl-X, Df(3R)Tl-P and Df(3R)ro80b and mutations in 97D are consistent with the results of Kidd et al. (1999) and Knibb et al. (1993).
Our studies mapped the taxi (tx) locus more precisely and provided information about its mutant phenotype and the strength of mutant alleles. tx maps to the region of overlap between the proximal ends of Df(3R)Tl-I and Df(3R)Tl-X and the distal end of Df(3R)Espl3 in proximal 97B. The phenotypes of tx1/Df individuals were more severe than the phenotypes of tx1 homozygotes, indicating that tx1 is hypomorphic. tx1 homozygotes frequently held only one wing out and rarely showed other wing phenotypes. tx1/Df individuals had strongly outheld wings that were thickened, slightly cupped and occasionally blistered. Df(3R)Espl3/Df(3R)Tl-X and Df(3R)Espl3/Df(3R)Tl-I exhibited a phenotype which probably represents the tx null phenotype. The thickened and opaque wings were always held out and somewhat crumpled when they were not filled with melanized fluid. Nevertheless, viability was only slightly reduced and females were fertile.
T(Y;3)G75, with a breakpoint in 97B, exhibited position effect suppression of both tx and scribbled (scrib). T(Y;3)G75 in combination with Df(3R)Espl3, Df(3R)Tl-X, Df(3R)Tl-P, Df(3R)Tl-I or Df(3R)ME61 showed a taxi phenotype slightly more severe than that of tx1/Df; most had strongly outspread wings with clear blisters. T(1;Y)G75/P{lacW}scribj7B3 showed a weak scrib phenotype: females laid eggs that did not hatch. Other T(Y;3) chromosomes with similar third chromosome breakpoints (T(Y;3)R87, T(Y;3)R71 and T(Y;3)B158) gave viable and visibly normal phenotypes in combination with Df(3R)ME61.
Our cytology of T(3;4)A30 places its third chromosome breakpoint at 97B9-C1. T(3;4)A30 is lethal in combination with deletions of 97BC. According to David Bilder, University of California at Berkeley (personal communication), T(3;4)A30 complemented scrib loss-of-function alleles, and rescuable maternal effect phenotypes of scrib were not rescued by paternal contribution of Ts(3Lt;4Rt)A30, i.e. the translocation segregant missing the distal portion of 3R could not provide scrib+ function to rescue the abnormal cuticular phenotypes of embryos derived from homozygous scrib maternal germ line clones when it was contributed by the male parent. This places the T(3;4)A30 breakpoint proximal to scrib. We found that T(Y;3)G75 may have position effects on the lethal gene(s) disrupted by T(3;4)A30, since T(Y;3)G75/T(3;4)A30 flies had reduced viability.
Acknowledgments: We thank Pier Paolo d’Avino his observations and David Bilder for sharing his data. This work was supported by NIH grant R24 RR14106 and NSF grant DBI-9816125 to K.R.C., MRC grant GB8225539 to Michael Ashburner and MRC grant G9901264 to David Glover.
References: Anderson, K.V. et al., 1985, Cell 42: 779-789; Adams, M.D. et al., 2000, Science 287: 2185-2195; Carfagna, M., and B. Nicoletti 1963, Dros. Inf. Serv. 38: 32; Gatti, M., and S. Pimpinelli 1983, Chromosoma 88: 349-373; Hartley, D.A. et al., 1988, Cell 55: 785-795; Karsch-Mizrachi, I., and S.R. Haynes 1993, Nucleic Acids Res. 21: 2229-2235; Kidd et al., 1999, Dros. Inf. Serv. 82: 114-115; Knibb, W.R. et al., 1993, Molec. Gen. Genet. 239: 109-114; Preiss, A. et al., 1988, EMBO J. 7: 3917-3927; Ranz, J.M. et al., 2001, Genome Res. 11: 230-239; Seattle-La Jolla Drosophila Laboratories, 1971, Dros. Inf. Serv. 47 (Suppl.); Spradling, A.C. et al., 1999, Genetics 153: 135-177; Tearle and Nusslein-Volhard, 1987, Dros. Inf. Serv. 66: 209-269.