Haywood, Annika F.M., Lisa D. Saunders, and Brian E. Staveley. 2002. Dopa decarboxylase(Ddc)-GAL4 dramatically reduces life span. Dros. Inf. Serv.
|
|
|
|||
|
|
||||
Dopa decarboxylase(Ddc)-GAL4 dramatically reduces life span.
Haywood, Annika F.M., Lisa D. Saunders, and Brian E. Staveley. Department of Biology, Memorial University of Newfoundland, St. John's, Newfoundland & Labrador, Canada A1B 3X9; telephone: (709) 737-4317, telefax: (709) 737-3018, bestave@mun.ca
Abstract
The
GAL4 system is extensively used for the directed expression of genes of interest
in Drosophila melanogaster. One main property of this system is that GAL4, the yeast transcription
factor, is believed to be inactive in Drosophila and that its expression under the control of D.
melanogaster-specific promoters will have
little effect on its own cells. We
have shown that expression of GAL4
under the control of the dopa decarboxylase (Ddc) promoter element does have an effect upon adult viability.
Although Ddc-GAL4 heterozygotes appear normal when raised at 25°C,
the homozygotes have a have a greatly reduced life span. While the homozygotes simply do not survive,
Ddc-GAL4 heterozygotes demonstrate normal
locomotion over their life span.
Introduction
The
GAL4 ectopic expression system (Brand and Perrimon, 1993; reviewed in Phelps
and Brand, 1998) has become an extremely useful approach for the study of
specific genes in Drosophila melanogaster. This expression system relies upon GAL4
to bind the upstream activation sequence (UAS) in order to activate transcription
of the target gene. A
variety of transgenic Drosophila lines are
readily available that express GAL4
in specific tissues or cell types. Our
laboratory is interested in models of neurodegenerative diseases and we have
initiated work with the dopa decarboxylase (Ddc)-GAL4 transgenic expression lines to model Parkinson’s disease in Drosophila.
The Drosophila melanogaster model of Parkinson’s disease was developed by the generation
of transgenic lines bearing wild type and mutant forms of the human a-synuclein gene cloned downstream of the UAS yeast promoter
(Feany and Bender, 2000). There
is no apparent Drosophila homologue
of a-synuclein, but expression of the human a-synuclein protein in the Drosophila nervous system recapitulated some features of Parkinson’s
disease. Expression of a-synuclein
in the dopaminergic neurons (Feany and Bender, 2000) was driven by a transgene
comprised of the dopa decarboxylase
(Ddc) gene promoter cloned upstream of GAL4. Originally,
this transgene was developed to examine a Drosophila model of cocaine addiction (Li et al., 2000). The
Parkinsonian flies, apparently normal at a young age, demonstrated a premature
loss of locomotor (climbing) ability, loss of the dopaminergic neurons and
accumulation of a-synuclein-containing
inclusions. In addition, GMR-GAL4 driven expression in the developing eye resulted in age-dependent
retinal neurodegeneration. Subsequent
treatment of the transgenic a-synuclein expressing flies with a number of pharmacological
agents such as the dopamine precursor L-DOPA, dopamine receptor agonists (bromocriptine,
pergolide and SK&F38393) and the anticholinergic atropine, all restored
or partially restored the age-dependent loss climbing ability (Pendleton et
al., 2002). Further, this model has been used to examine
the suppression of the a-synuclein toxicity by the molecular chaperone, hsp70 (Auluck
et al., 2002). A good understanding of the effects of
Ddc-GAL4 expression in Drosophila melanogaster is essential to properly interpreting this model system.
As
a prelude to our exploration of models of Parkinson's disease in Drosophila, we began to investigate the biological properties
of the Ddc-GAL4 driver lines.
We have shown that Ddc-GAL4 causes
reduced viability and is therefore not inactive in D. melanogaster.
Materials and Methods
Fly stocks and culture: Ddc-GAL4 4.3D and Ddc-GAL4 4.36 flies (Li et al., 2000) were obtained from Jay Hirsh at the Department of Biology, University of Virginia and w1118 was obtained from Dr. Howard Lipshitz at the Hospital for Sick Children in Toronto. To obtain heterozygotes, Ddc-GAL4 homozygous males were crossed to w1118 females. All flies were cultured on standard cornmeal/yeast/molasses/agar media at 25°C.
Aging analysis: Adult males were aged in small groups upon standard media at 25°C and scored for viability every two to three days as described previously (Staveley et al., 1990). The number of individuals aged are as follows: Ddc-GAL4 4.3D homozygotes n = 102; Ddc-GAL4 4.36 homozygotes n = 133; Ddc-GAL4 4.3D heterozygotes n = 280; Ddc-GAL4 4.36 heterozygotes n = 119; w1118 n = 83).
|
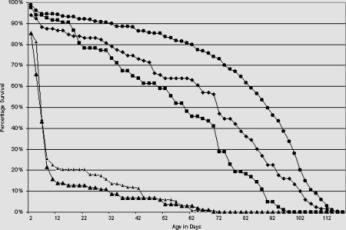
Figure 1. Ddc-GAL4 reduces life span. Male homozygotes of the independent insertions of the Ddc-GAL4 transgene, Ddc-GAL4 4.3D (large triangles) and Ddc-GAL4 4.36 (small triangles) both display a greatly reduced life span. The heterozygotes of Ddc-GAL4 4.3D (solid circles) and Ddc-GAL4 4.36 (solid diamonds), however display a normal, if not extended life span when compared to the control w1118 (solid squares) individuals. The number of males aged are as follows: Ddc-GAL4 4.3D homozygotes, n = 102; Ddc-GAL4 4.36 homozygotes, n = 133; Ddc-GAL44.3D heterozygotes, n = 280; Ddc-GAL4 4.36 heterozygotes, n = 119; w1118, n = 83.
|
Locomotion
assay: The flies were assayed for their
ability to climb in a manner similar to that described by Feany and Bender
(2000). Every 4 to 5 days, 10
male flies of a cohort of aged flies were assayed for their ability to climb
to the top of the vial within a period of 18 seconds. Twenty trials were carried out for each time point.
Results and Discussion
An investigation of the baseline biological consequences of GAL4 expression as directed by the dopa decarboxy-lase(Ddc) promoter is essential to our studies of Parkinson's disease models in Drosophila. We began by crossing the Ddc-GAL4 4.3D and Ddc-GAL44.36 driver lines to w1118 to conduct climbing assays. It quickly became apparent that the stocks of both Ddc-GAL4 insertion lines required extra care to maintain and that the flies were apparently short-lived in both cases. As a result we decided to conduct longevity trials.
|
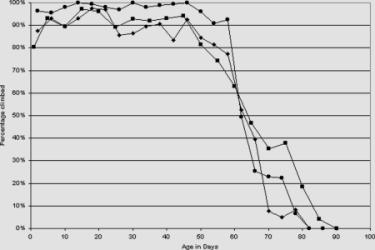
Figure 2. Ddc-GAL4 heterozygotes have normal locomotor (climbing) activity.
The heterozygotes of Ddc-GAL4 4.3D (solid circles) and Ddc-GAL44.36 (solid diamonds) display a normal
level of climbing ability when compared to the control w1118 (solid squares) flies. The locomotion of homozygotes was not
measured due to poor viability. |
Homozygotes of both insertions of the Ddc-GAL4 trans-gene display a greatly reduced life span (Figure 1). For example, by the 6th day, Ddc-GAL44.3D and Ddc-GAL44.36 homozygous males showed only 43% and 44% survival, respectively. While 50% of Ddc-GAL44.3D male het-erozygotes survive past the age of 88 days and Ddc-GAL44.36 male heter-ozygotes past 70 days of age. We assigned w1118 as the control strain in these experiments, 50% of which survived between 58 and 60 days. The shorter median life span of the control may reflect an insufficiency of the w1118 stock rather than an increase in viability the heterozygotes. It is important to note that the very similar longevity profile of the two independent insertions of Ddc-GAL4 suggest that the reduction in life span is due to the expression of the transgene and not the site of insertion.
The reduced viability of the Ddc-GAL4 homozygotes forced us to reexamine the locomotor activity of the Ddc-GAL4 heterozygotes (Figure 2). However, the Ddc-GAL4 4.3D and Ddc-GAL4 4.36 heterozygotes retain their ability to climb with age in a manner similar to the w1118 controls. Due to greatly reduced viability, the locomotion of homozygotes was not measured.
The UAS/GAL4 ectopic expression system has made it possible to express genes and test the effects of overexpression. The Ddc-GAL4 driver has been used for expression of transgenes in the dopaminergic neurons including expressing the a-synuclein gene to model Parkinson’s disease (Feany and Bender, 2000). Although no UAS (cggagtactgtcctcc) promoter sequences are found in D. melanogaster (Berkeley Drosophila Genome Project, pers. comm.), our laboratory has demonstrated that expression of GAL4 in the eye with the GMR-GAL4 transgene leads to increased levels of apoptosis and morphological defects (Kramer and Staveley, submitted). Although the mechanism by which GAL4 induces cell death is unclear, death of the dopaminergic neurons could certainly result in premature lethality in these flies.
Acknowledgments: This work was funded by the Natural Sciences and Engineering Research Council of Canada and the Dean of Science of Memorial University of Newfoundland (start-up funds) to B.E.S. L.D.S. was funded by an Undergraduate Student Research Award from the Natural Sciences and Engineering Research Council of Canada. We thank Lloyd Smith for design and production of the climbing assay device. We thank Dr. Jay Hirsh at the Department of Biology, University of Virginia, and Dr. Howard Lipshitz at the Hospital for Sick Children in Toronto for providing fly stocks.
References: Auluck, P.K., H.Y.E. Chan, J.Q. Trojanowski, V.M.-Y. Lee, and N.M. Bonini 2002, Science 295: 865-868; Brand, A. H., and N. Perrimon, 1993, Development 118: 401-415; Feany, M.B., and W.W. Bender 2000, Nature 404: 394-398; Li, H., S. Chaney, I.J. Roberts, M. Forte, and J. Hirsh 2000, Curr. Biol. 10: 211-214; Pendleton, R.G., F. Parvez, M. Sayed, and R. Hillman 2002, J. Pharmacol. Exp. Ther. 300: 91-96; Phelps, C. B., and A.H., Brand 1998, Methods 14: 367-379; Staveley, B.E., J.P. Phillips, and A.J. Hilliker 1990, Genome 33: 867-872.