Ozsoy, E.D., and A.N. Bozcuk. 2000. The P-M characteristics of some natural Drosophila melanogaster populations from Turkey. Dros. Inf. Serv. 85: 59-62.
|
|
|
|||
|
|
||||
The P-M characteristics of some natural Drosophila melanogaster populations from Turkey.
Ozsoy, E.D., and A.N. Bozcuk. Department of Biology, Hacettepe University, 06532 Beytepe, Turkey.
The number of P transposable elements in any natural population of Drosophila melanogaster can categorically be characterized within the dysgenesis system, P and M (Kidwell,1986; Engels, 1996). In this system, P strains are the ones that have many P elements with the potential of strong activity (i.e., activity by a full-sized element to induce dysgenesis), and with a strong repression of that activity. Strains classified as Q are a subset of P strains which consist of P elements of lesser activity but of strong repression ability. Finally, M strains are the ones without strong activity potential and strong repression, corresponding to their complete lack of any P element (Kidwell, 1986). A subcategory of M, termed M’, also exists with considerable variation in P activity regulation (Anxolabéhére et al., 1985). The P element-associated dysgenesis, hybrid dysgenesis, of an individual fly is thought to be determined by the very presence of genomic P elements in that fly, and the categories are constructed by the levels of dysgenesis measured by appropriate genetical tests (Kidwell, 1986; Engels, 1996). Geographical variation in P element numbers of natural D. melanogaster populations (corresponding to their P-M status) has been documented. The picture is that of considerable variation within populations, but of similarity across populations over large, continental scales (Anxolabèhère et al., 1988). In particular, lines from North America have many full-size P elements, with smaller nonautonomous deletion derivatives. European lines have relatively few full-size, especially in Mediterranean, but with many copies of a short derivative, KP element (Anxolabèhère et al., 1985, 1988; Black et al., 1987). The numbers of all-size P elements declines from west to east in Central Asia, but all copy numbers rise high again in eastern China and Japan (Todo et al., 1984; Sakoyama et al., 1985; Anxolabèhère et al., 1988, 1990). The situation is complicated for Australia; samples collected some 20 years ago in eastern Australia showed a strong clinal P-M pattern, populations from the north being the strong P, and the populations to the south grading from Q to M (Boussy, 1987; Boussy and Kidwell, 1987; Boussy et al., 1988). Sampling approximately 10 years later revealed much decay in this Australian pattern. The clinality was much reduced in amplitude, the northernmost populations being not the previous extreme P populations (Boussy et al., 1998; French et al., 1999).
In this study we sampled 5 geographically different natural D. melanogaster populations from Turkey and determined their P-M statuses. Three Eastern Mediterranean (Yumurtalik, Erzin, Payas) and two central Anatolian (Eryaman, Cebeci; two local populations with a distance of 20 km from each other in Ankara) populations were collected during the summer of 1995. At each location the number of the flies collected exceeded 30 individuals per population. Temperature dependent sterility trait (Gonadal dysgenesis, GD) of hybrid dysgenesis was used to construct P-M categories. The gonadal dysgenesis assay allows one to analyze the results of dysgenic crosses with marker strains, the visible changes in the overall morphology of gonads of the F1 female progenies caused by P mobility at developmental temperature above 25°C (exactly 29°C in this study), to determine P-M categories in an easiest way. By the diagnostic cross A, it is possible to measure the potential of a strain to induce gonadal dysgenesis trait. The cross A* allows one to determine the regulatory ability (i.e., suppression) of a strain (see Kidwell, 1986, for experimental setup and the marker strains used in these specific crosses). Assays were performed simultaneously in 2nd generation for all the populations maintained in laboratory conditions after collection. At least 50 F1 females were dissected per cross per population to decide the P activities and the regulatory potentials of those activities.
|
Table 1.
The P-M statuses of different geographical D. melanogaster strains determined by gonadal
dysgenesis (GD) of hybrid dysgenesis.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 1 shows the percentage dysgenesis levels per population
for the activity potentials and regulatory abilities. Almost all populations
are M’ strains on the basis of the criteria defined in Kidwell (1986).
The only population to which a different category could be assigned, namely
Q, is Yumurtalik, a Mediterranean one. A curious observation is that for one
of the strains, Eryaman, the decisiveness of the GD determination from its
A* cross is not unambiguous as for the rest of the populations; control (25°C)
|
|
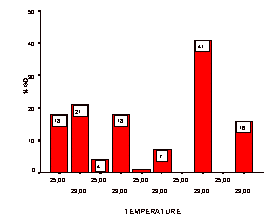
Figure 1. Dysgenesis levels of the strains in A crosses. Strains are given from left to right in their respective GD values as denoted in Table 1. |
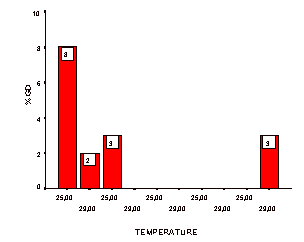
We think that this might be due to differential regulatory response to developmental temperature in this strain for the dysgenesis trait. Another feature of our data is the distinct variability of the M’ populations, as determined by GD values from the respective A* crosses (Table 1, Figure 2). This is the typical M’ regulation documented for other strains, too (Anxolabéhére et al., 1985). Population specific developmental responses to different temperatures in this study are illustrated in Figures 1 and 2.
The populations were sampled from two ecologically distinct regions; Mediterranean and central Anatolian parts of Turkey, the latter being characterized of a typical land climate. Overall, the P-M status distribution of the populations is monotonic, almost all being M’, irrespective of geographical origin. This picture is quite similar to that found for Eurasian strains (Periquet et al., 1989). However, taken together with those of the only other report from Turkey by Konac and Bozcuk (1990) of a more southwardly Q strain, our results are not completely satisfactory to draw a general conclusion for Turkey concerning P-M status.
|
|
Figure 2. Dysgenesis levels of the strains in
A* crosses. Strains are
given from left to right in their respective GD values as denoted in
Table 1. |
We conclude that the correspondence of our results to the gross Eurasian pattern could support the hypothesis that the evolution of the P-M status is taking place at a low rate across broad Eurasian geography. We also point to the requirement of a molecular analysis for further elaboration of the P-M categories we have constructed here.
Acknowledgment: We thank Dr. M.G. Kidwell for providing the marker strains.
References: Anxolabéhére, D., D. Nouaud, G. Periquet, and P. Tchen 1985, Proc. Natl. Acad. Sci. USA 82: 5418-5422; Anxolabéhére, D., M.G. Kidwell, and G. Periquet 1988, Mol. Biol. Evol. 5: 252-269; Anxolabéhére, D., K. Hu, D. Nouaud, and G. Periquet 1990, Gen. Sel. Evol. 16: 15-26; Black, D.M., M.S. Jackson, M.G. Kidwell, and G.A. Dover 1987, EMBO J. 6: 4125-4135; Boussy, I.A., 1987, Gen. Res. 49: 11-18; Boussy, I.A., and M.G. Kidwell 1987, Genetics 115: 737-745; Boussy, I.A., M.J. Healy, J.G. Oakeshott, and M.G. Kidwell 1988, Genetics 119: 608-615; Boussy, I.A., M. Itoh, D. Rand, and R.C. Woodruff 1998, Genetica 104: 45-57; Engels, W.R., 1996, In: Transposable Elements, (Saedler, H., and A. Gierl, eds.), 103-123, Springer-Verlag, Berlin Heilderberg; French, D.J., R. Corish, M. Shi, and G.A. Dover 1999, Gen. Res. 73: 205-216; Kidwell, M.G., 1986, In: Drosophila, A Practical Approach (Roberts, D.B., ed.), 59-81, IRL Press, Oxford; Konac, T., and A.N. Bozcuk 1990, Tr. J. of Biology 14: 32-41 (in Turkish); Periquet, G., S. Ronsseray, and M.H. Hamelin 1989, Heredity 63: 47-58; Sakoyama, Y., T. Todo, S. Ishiwa-Chigusa, T. Honjo, and S. Kondo 1985, Proc. Natl. Acad. Sci., USA 82: 6236-6239; Todo, T., Y. Sakoyama, S.I. Chigusa, A. Fukunaga, T. Honjo, and S. Kondo 1984, Jpn. J. Gen. 59: 441-451.