| BEN |
| BOTANICAL ELECTRONIC NEWS |
|---|
| ISSN 1188-603X |
|---|
Dr. A. Ceska, P.O.Box 8546, Victoria, B.C. Canada V8W 3S2
A RADICAL VIEW OF BRYOPHYTE BIOLOGY: MOSSES ARE FROM MARS, VASCULAR PLANTS ARE FROM VENUS
From: Brent Mishler [bmishler@socrates.berkeley.edu]
In what ways does bryophyte biology differ from that of vascular plants (tracheophytes)? The short answer: in almost every way possible! The groups didn't evolve on different planets, but their differences could almost make you think they did. They certainly adopted very different approaches to being a land plant on this planet. Many aspects need much more study, but what is known about bryophyte biology suggests that in general the bryophytes differ in most ways in their biology, ecology, and evolution from tracheophytes.
Major differences in bryophyte biology from tracheophytes include:
- Haploid dominance in the alternation of generations.
The green, vegetative part of the life-cycle in bryophytes is haploid. Without the genetic benefits of dominance, genes acting in the gametophyte are presumably subject to relatively severe selection.
- Extensive phenotypic plasticity.
Studies have shown that bryophytes tend to have very high amounts of morphological and physiological plasticity. This may compensate for their demonstrated low levels of ecotypic differentiation (perhaps due to haploidy).
- Poikilohydry and desiccation-tolerance.
Poikilohydry is the rapid equilibration of the plant's water content to that of the surrounding environment, while desiccation tolerance is the ability of a plant to recover after being air-dry at the cellular level. All bryophytes have these abilities to some extent, but this was lost in the larger, more complex, and endohydric tracheophytes.
- Need for free water for sexual reproduction.
A residual feature of the early land plants is the constraint imposed by the swimming sperm. Swimming gametes have short dispersal distances which leads to frequent inbreeding in monoicous species and lack of sporophyte production in dioicous species.
- The clump as a "super-organism".
Many mosses and some liverworts are essentially social organisms. This results from the combination of clonal growth, poikilohydry, and external water conduction. The plants in a clump are subject to natural selection as a group. Intimate contact of each vegetative cell with the environment, due to poikilohydry, lends itself to interplant chemical communication via pheromones.
- Heavy reliance on asexual reproduction.
Due to the difficulty of achieving fertilization, many bryophytes have evolutionarily lost functional sexuality. Since bryophytes grow from an apical cell, somatic mutation allows genetic variation even within clones.
- Small stature and the occupation of microhabitats.
Small size, lack of roots, and poikilohydry means that bryophytes are in a close relationship with only their immediate microenvironment. Over geological time, they may be less influenced by climatic change, and linger in refugial habitats.
- Less selection pressure from the biotic component of the environment than from the physical component.
Vagility and establishment abilities of bryophytes are relatively poor. Available substrates are not filled in most mesic and xeric environments (although they may be in some hydric environments). The presence of other bryophytes nearby often appears beneficial to growth.
- Relatively slow evolutionary rates in morphology.
The fossil record of bryophytes indicates that ancient forms are very similar to modern ones. Biogeographically, bryophytes tend to follow the same historical patterns of disjunction as tracheophytes, but at a lower taxonomic level. This may indicate that developmental constraints play an unusually important role.
The overall effect of these features on the evolutionary ecology of bryophytes makes them profoundly different. By studying bryophytes and comparing their life-style to that of tracheophytes, we can learn to observe structure closely, think critically about evolutionary inferences, and comprehend how different lineages can take different functional paths in response to the same stimuli.
[Abridged from Mishler, B.D. 2001. "Book review - The biology of bryophytes: Bryophytes aren't just small tracheophytes." American Journal of Botany 88: 2129-2131. - A review of: Bryophyte Biology. by A. Jonathan Shaw and Bernard Goffinet (eds.). 2000. Cambridge University Press. x + 476 p. - For the review of this book by Wilf Schofield see also BEN # 259.]
BOOK REVIEWS AND NOTICES
From: Adolf Ceska [aceska@victoria.tc.ca]
- Jacobsen, Arthur Lee. 2001.
- Wild plants of Greater Seattle: A field guide to native and naturalized plants of the Seattle area. Arthur Lee Jacobson, Seattle. 494 p. ISBN 0-9622918-2-x [softcover] Price: US$24.95
- Available from:
-
Arthur Lee Jacobson
2215 E. Howe Street
Seattle, WA 98112
e-mail: alj@consultant.com
web site: http://www.arthurleej.com/
Line drawings (most of them by J.R. Janish from Hitchcock et al. Vascular plants of the Pacific Northwest) accompany descriptions of about 500 vascular plants. The descriptions contain information about the main diagnostic characters, intraspecific variation, biology and ethnobotany of each species. Forty introductory pages list plants of various habitats, summarize their edibility and give month-by-month accounts of their phenology. An annotated checklist of about 1,270 species gives more details on their distribution in the Greater Seattle area.
- Kershaw, Linda, J. Gould, D. Johnson, & J. Lancaster [eds.]. 2001.
- Rare vascular plants of Alberta. University of Alberta Press & The Canadian Forest Service, Edmonton, Alberta. xliv+484 p. ISBN 0-88864-319-5 [softcover] Price: CDN$29.95
- Available from:
-
Raincoast Books
9050 Shaughnessy Street
Vancouver, BC, CANADA V6P 6E5
Customer service: 1-800-511-6024 Fax: 1-800-565-3770
E-mail: custserv@raincoast.com or info@raincoast.com
Internet: http://www.raincoast.com/
This publication describes approximately 485 vascular plants that are considered rare in Alberta. In a one-species-per-page format, this manual gives the name of the species, most common synonyms, description, habitat and notes on similar species. Species are illustrated with colour photographs and/or line drawings, and for most species, maps of their distribution in North America and in Alberta are provided. The work is a result of intensive floristic surveys that included 30 direct contributors and about 100 amateur botanists.
Besides the species treatments, the book gives a good introduction to problems of rare plant studies, and a good description of the phytogeography of Alberta. Several appendices provide interesting reading (yet another key to Alberta Botrychium species!), and an illustrated glossary and close to 300 literature citations conclude this handy publication.
When I first opened the book, I had the impression that it was yet another book in the successful plant guide series published by Lone Pine Publishing. Not so. However, although it was published by the University of Alberta, editor Linda Kershaw, formerly of Lone Pine, had an obvious and significant influence on the format of this publication. The University of Alberta Press, Canadian Forest Service, Alberta Native Plant Council (Alberta's own version of a Native Plant Society), four editors, 30 contributors and about 100 volunteers should be commended for this great book.
- Phipps, James B. & Paul M. Catling. [Eds.] 2001.
- Bioconservation and systematics: proceedings of the Canadian Botanical Association Conference Symposium in London, Ontario, June 2000. Canadian Botanical Association, Ottawa. iii+101 p. ISBN 0-9689565-0-5 [softcover] Price: CDN$23.00 (US$17.00) in Canada and USA, CDN$33.00 (US$21.00) overseas.
- Ordering information:
-
Send a check made out to "Canadian Botanical Association" to:
Dr. Mel Fisher
Box 160
407 Main Street
Aberdeen, Saskatchewan
Canada S0K 0A0
Prices include shipping and handling.
This is a collection of papers presented at the "Bioconservation and Systematics" symposium organized by the Canadian Botanical Association (CBA/ABC) at the annual general meeting in 2000. The symposium addressed the theme common to other similar symposia, such as one organized by the Missouri Botanical Garden (Annals of the Missouri Botanical Garden, vol. 87, no. 1, 2000) or by the Jepson Herbarium (Madroño, vol. 47, no. 2, 2000): What is the role of biosystematics in the exploration and preservation of plant biodiversity?
The symposium organized by the Canadian Botanical Association included the following contributions:
- Bioconservation and Systematics - Symposium Introduction (James Phipps)
- A never-ending role for biosystematics in the protection of vascular plant biodiversity in Canada (Paul Catling)
- Planning with plants in Illinois (Kenneth Robertson)
- The role of Conservation Data Centres in the conservation of Canada's flora (Michael Oldham & Peter Sorrill)
- Floristics and conservation: an example from Newfoundland (Luc Brouillet)
- Evaluating hybridization as a cause for species endangerment: a role for systematics in plant conservation (Brian Husband & Kevin Burgess)
- Can conservation help conserve rare plants in the twenty-first century? (Anton Reznicek)
PALINDROMES REVISITED
My posting on palindromes stimulated more "Letters to the editor" than any other botanical posting in BEN. - AC
Costas A. Thanos [cthanos@biol.uoa.gr] wrote:
In regard to the palindromic phrase in BEN 279, a few comments:
- The phrase was 'invented' by Byzantine Greeks (the 'middle' ones, not ancient Greeks) - so it was an early Christian phrase.
- The original inscription was made on a fountain outside Saint Sophia, in Constantinople (today Istanbul, Turkey), the capital of the Byzantine Empire. It was made during the 6th century AD, during the reign of Emperor Justinianus (who commanded the construction of the magnificent cathedral).
- The correct translation is: 'Wash sins not only face'.
The Greek word (although palindrome is perfectly Greek as well) for the phrases that can be read both ways is 'cancer phrase' or 'cancer text'. Cancer is crab of course.
The spelling in Greek is [top is original text version and bottom is graphic]
Î^ÝÎ^ŮΨÎ^ßÎ^Ý Î^ŃÎ^ÝÎ^ßÎ^ÜÎ^×Î^ÜÎ^ŃΤÎ^Ń Î^ÜÎ^× Î^ÜÎ^ßÎ^ÝÎ^ŃÎ^Ý Î^ßΨÎ^ŮÎ^Ý
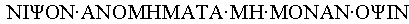
- [For the text version,] I guess you will need to install Greek fonts to read it properly. The transliteration you used in the BEN is the closest possible phonetically.
Subscriptions:Send "subscribe BEN-L" or "unsubscribe BEN-L" (no apostrophes) to majordomo@victoria.tc.ca.
Send submissions to BEN-L@victoria.tc.ca
BEN is archived at: http://www.ou.edu/cas/botany-micro/ben/
http://www.ou.edu/cas/botany-micro/ben/ben280.html