Flores, S.V., P. Iturra, and R. Godoy-Herrera. 2001. Genetic differentiation between three populations of Drosophila pavani that live in different breeding sites in nature. Dros. Inf. Serv. 84: 67-70.
|
|
|
|||
|
|
||||
Genetic differentiation between three populations of Drosophila pavani that live in different breeding sites in nature.
Flores, S.V., P. Iturra,
and R. Godoy-Herrera. Programa de Genética Humana, Instituto
de Ciencias Biomédicas, Facultad de Medicina, Universidad de Chile.
email: sflores@machi.med.uchile.cl
Natural selection and genetic drift acting on populations of one species which exploit distinct resources originate genetic differences which may lead to reproductive isolation between them (Ridley, 1996). This is the first step in formation of new species (Coyne and Orr, 1998). Drosophila pavani is endemic to Chile, even if it has recently also been collected in Ecuador (Vela and Rafael, 2001). In Chile, D. pavani has been found to be an unified species, that is not divided into subspecies (Brncic, 1970). On the other hand, very little is known about the ecology of D. pavani. However, we have found that D. pavani can use as breeding sites a diversity of substrates which differ in several ecological features. Thus, Manriquez and Benado (1994) reported that the cactus Echinopsis chilensis, which may produce substances as alcaloids and triterpens (see Barker, 1990), is an endemic breeding site for D. pavani. Adults of the species also may emerge from overipe apples fallen on the ground. In this type of fruits, concentration of ethanol and acetic acid is high (Parsons, 1983). Thus, D. pavani represents a favorable material for study of reproductive isolation between populations which exploit different ecological resources. The purpose of this work is to investigate genetic differentiation between three geographically different populations of D. pavani with particular reference to intercrosses between them and molecular genetic markers. One population was formed with ancestors which had emerged from overipe tissue of E. chilensis growing in Til-Til, a dry place at 50 Km Northwest from Santiago (the Til-Til strain) and the other two strains were originated from ancestors which emerged from overipe apples collected in Chillán, 420 Km South away from Santiago (the Chillán strain), and in La Florida located in Santiago itself (the La Florida strain).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The stocks
are kept by mass culture in the Human Genetic Program, Faculty of Medicine,
University of Chile. They were
all reared in a constant environment under constant light at 18°C.
Crosses: Twenty-day-old virgin males and females of the Til-Til, Chillán and La Florida strains were reciprocally crossed. The F2 and backcross generations were also obtained (Table 1). Homogametic matings within each strain were also performed to serve as control for the other crosses (see Table 1). After that, females of each cross were allowed to lay eggs for 4-5 days in half-pint bottles containing nutritive medium (Burdick, 1954). Previously, 3-4 drops of live yeast cream were deposited onto the surface of the medium. After 18-20 days the number of males and females emerged from each type of cross was recorded.
DNA Genetics Markers: Six larval genotypes of the Chillán strain and two other groups of seven of the Til-Til and La Florida strains were, respectively, used in this work. Before all DNA extractions, the specimens were individually cleaned and washed in sterile destilled water. Each larva was smashed in 200 ml of Chelex 5%. RAPD analysis was carried out using 8-mer oligonucleotide primers from Operon Technologies Inc: OP-G11, OP-M13, OP-A20, OP-G18 and OP-P04. The conditions of the PCR reactions were the same described by Iturra et al. (1998). After each PCR reaction, aliquots of amplified DNA from each of the individuals tested were electrophoresed on agarose gel containing 0.5% TBE. The individual samples obtained with each one of the primers were run together with 1 ml of DNA molecular weight marker to determine the approximate length of the PCR products. After that, we used the pattern bands of each of the larval genotypes to build individual matrices scoring presence (1) or absence (0) of each band. Genetic distances between individuals were estimated by using the Apostol (1994) and Sokal and Sneath (1963) methods included in the RAPD-Distance software. The agreement between both methods was estimated by the Mantel test. The corresponding dendrograms (see one of them in Figure 2) were also yielded by using the NTSYS-pc software (Rohlf, 1994).
Table 1 shows that Til-Til females crossed with males of the Chillán and La Florida strains do not produce offspring. However, the corresponding reciprocal crosses (Til-Til males ´ Chillán females, and Til-Til males ´ La Florida females) originate a substantial number of F1 adult flies of both sexes (Table 1). In contrast, the two reciprocal crosses between the Chillán and La Florida strains produce abundant F1 males and females. It is also interesting to note that all F1 flies produce abundant F2 fertile adult flies (Table 1). However, the F1 males, obtained to cross the Chillán and Til-Til strains, backcrossed with Chillán females do not produce offspring (Table 1). In contrast, the F1 females crossed with Chillán males originate adult flies of both sexes. Finally, the reciprocal backcrosses between the F1 and the Til-Til strain produce abundant number of males and females (Table 1).
|
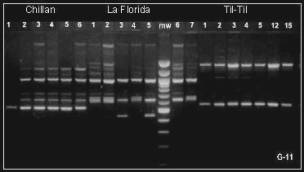
Figure 1. Gel electrophoresis of RAPD products by using G-11 primer.
Figure shows the migration patterns of DNA bands of six genotypes
of the Chillán strain, and samples of seven genotypes obtained,
respectively, from the La Florida and Til-Til strains. (mw: molecular
weight marker). |
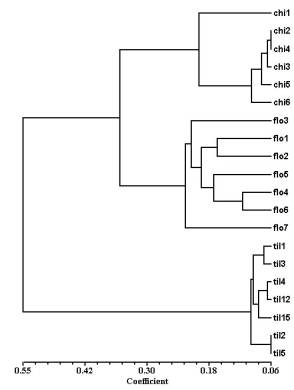
Dendrogram (see Figure 2) obtained by using the UPGMA method (Rohlf, 1994) is in good agreement with Figure 1, that is the Chillán and La Florida populations yield very close clusters clearly different to the cluster of the Til-Til population (Figure 2). The mean of genetic distance between individuals within each of the samples is: i) the Chillán strain, 0.13 ± 0.07, ii) the La Florida strain, 0.20 ± 0.01, and iii) the Til-Til strain, 0.09 ± 0.01. These findings suggest that the sample of Til-Til flies is genetically more homogeneous that those of the Chillán and La Florida strains.
The results
of this work show that flies of the Til-Til population of D. pavani are in part genetically isolated from the Chillán
and La Florida populations. These last two populations are separated by 420 Km, but the
results (Table 1) suggest that there is not reproductive isolation between
them. On the other hand, the
La Florida and Chillán strains were formed with ancestors which had
emerged from rotten apples. In
contrast, the Til-Til strain was originated with flies emerged from overipe
tissue of columnar cactus E. chilensis. We would like to suggest that the observed
reproductive isolation may be a consequence of genetic differentiation between
the examined D. pavani populations
due to the process of adaptation to very different breeding sites. However, we have not investigated whether
the observed restrictions in genetic flow are due to premating isolation or
postzygotic reproductive isolation.
|
Figure 2. Dendrogram showing the relationships
between samples of individuals of the Chillán, La Florida and
Til-Til strain (see text). The
number of larval genotypes tested were, respectively: i) the Chillán (chi) strain, 6; the La Florida (flo)
strain, 7, and iii) the Til-Til (til) strain, 7. |
It is interesting to note that the results yielded with the genetic markers are in good agreement with the hypothesis of adaptation of the Til-Til population to a breeding site substantially different to those of the Chillán and La Florida strains. That is, the band patterns and cladogram (Figures 1 and 2) indicate that the Til-Til population is genetically very different to the Chillán and La Florida strains. Taken together, our findings suggest that the populations of D. pavani here studied have accumulated genetic differences expressed in partial reproductive isolation between the Til-Til and the Chillán and La Florida strains. The isolation could have built because the populations have climbed separate adaptative peaks that correspond to distinct ecological niches, as suggested by
the substrates used as breeding sites. We especulate that the Til-Til ´ Chillán, and Til-Til ´ La Florida hybrids could show intermediate phenotypes between the parental strains and thus they may removed by selection. We are planning to investigate this further.
Acknowledgments: S.F. was supported by a fellowship from the ICBM (Instituto de Ciencias Biomédicas), Facultad de Medicina, Universidad de Chile.
References: Burdick, A.B., 1954, Dros. Inf. Serv. 28: 170; Apostol, B., P. Smouse, and J. Quattro
1992, Gen. Soc. Amer. 131: 479-491;
Barker, J., 1990, In: Ecological and Evolutionary Genetics
of Drosophila. (Barker, J.S.F., W.T. Starmer, and R. McIntre, eds). pp. 161-175,
Plenum Press, New York; Brncic,
D., 1970, In: Essays in Evolution and Genetics in Honor of Theodosius
Dobzhansky. (Hecht, M.K., and W.E. Steere, eds.).
pp. 401-436. Crofts, New York;
Coyne, J., and A. Orr 1998, Phil. Trans. R. Soc. Lond. B 353: 287-305;
Iturra, P., J.F. Medrano, M. Bagley, N. Lam, N. Vergara, and J.C. Marín
1998, Genetica 101: 209-213; Manriquez, G., and M. Benado 1994, Revista Chilena de Entomología
21: 185-186; Parsons,
P.A., 1982, Evol. Biol. 14: 297-328;
Rafael, V., and D. Vela 2001, Dros. Inf. Serv. 83: 85-88; Ridley, M., 1996, Evolution (Second edition). Blackwell Science, Oxford; Rohlf, F., 1994, NTsys-pc: Numerical taxonomy and multivariate
analysis system, Windows version 2.10ª, Applied Biostatistics Inc., New
York; Sokal, R., and P. Sneath
1963, Principles of Numerical Taxonomy. p. 128, Second Edition.