Kulkarni, Gauri V., Dileep N. Deobagkar, and Deepti D. Deobagkar. 2002. Hexokinase like protein from Drosophila melanogaster. Dros. Inf. Serv. 85: 65-68.
|
|
|
|||
Hexokinase like
protein from Drosophila melanogaster.
Kulkarni, Gauri V., Dileep N. Deobagkar, and Deepti D. Deobagkar. Molecular Biology Research Laboratory, Department of Zoology, University of Pune, Ganeshkhind, Pune-411007. Maharashtra, INDIA.
Summary
A genomic clone from D. melanogaster, when completely sequenced and analyzed, showed the presence
of an uninterrupted coding sequence of 1398 bp coding for a 465 amino acid ORF. Although this protein showed
overall 31% identity and 56% similarity to the mammalian HK1, critical glucose
binding residues are altered in this protein. In mice, a moonlighting role
has been assigned to HK1 testis specific isoform in the sperm egg interaction
suggesting that the glucose binding ability of hexokinases can be co-opted
for carbohydrate binding in the extracellular matrix of the egg. Conservation
of such homologue in Drosophila raises very interesting possibilities about moonlighting
roles of hexokinases during evolution.
Hexokinases ({ATP:d-hexose6-phosphotransferase}, EC2.7.1.1) are ubiquitous housekeeping enzymes catalyzing
the phosphorylation of glucose
by ATP, which occurs in all eukaryotic and prokaryotic cells as the first
step in the utilization of glucose (Grossbard and Schimke, 1966; Knutsen et
al., 1969; Faulkner and Jones, 1976; Peters and Neet, 1977).
Mammals show four hexokinase isoforms (Type I-IV) (Grossbard and Schimke,
1966). Apart
from acting as a hexokinase, multiple moonlighting roles have been assigned
to hexokinases. Rat brain HK-1 was shown to have autophosphorylating and protein
kinase activity (Adams et al.,
1994), the yeast hexokinase pII was shown to involved in repression
of SUC-II gene, which encodes for an enzyme involved in disaccharide metabolism
(Piller et al., 1998). Immunolocalization studies indicated the presence
of testes specific HK-1 isoform on the mouse sperm surface suggesting its
probable role in sperm and egg zona pellucida (ZP3) interaction (Kalab et
al., 1994; Travis et al., 1998). Based on biochemical and genetic analysis, four
isoforms of hexokinase from D. melanogaster have been reported (Murray and Ball, 1967; Cavener,
1980; Madhavan et al., 1972; Voelker
et al., 1978). Using
native gel electrophoresis, Murray and Ball (1967) have described a testes
specific isoform Hex T. Hex A and Hex B isozymes are products of a single
X-linked locus (8D/ X), which are largely expressed in adult thorax and larval
muscle tissue, respectively, while Hex C (52 E/ 2R) enzyme is expressed in
adult and larval fat body. Here,
we report the sequence analysis of a 465 amino acid coding ORF, which, although
it has homology to hexokinases, shows remarkable alteration in typical substrate
binding domains.
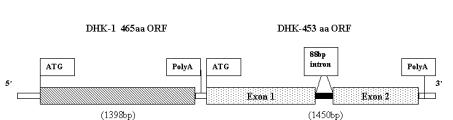
A genomic clone from D. melanogaster genomic library EMBL3 was completely sequenced. Analysis of a 4509 bp region revealed presence of two genes separated by a short variable-length intergenic sequence of 41 bp (Figure 1). The first gene exhibits an uninterrupted coding sequence of 1398 bp, while the second gene contains a coding sequence of 1359 bp that is interrupted by a single intron of 88 bp. The sequence is deposited in EMBL database (Accession No. AJ 271350). This sequence was localized to 97B on the third polytene chromosome (Data not shown), which is consistent with its localization reported in Drosophila Genome Project (Gadfly Gene CG54430) (Adams et al., 2000) and is different from that of HexA (8D/X) and HexC (51B/2R).
|
|
Further, glucose and ATP binding domains of DHK-465 and DHK-453 were compared with Drosophila hexokinases (Hex A and Hex C), mammalian and yeast hexokinase sequences (Figure 2A and B). DHK-465 sequence shows the presence of glucose binding domain at 138-165 aa and ATP binding domain at 68-100 aa residues, respectively. In DHK-453, glucose binding domain resides at amino acid position 133-161 and ATP binding domain resides at position 63-83, respectively. 8 major residues in the glucose binding domain are altered in DHK-465 protein (Figure 2A). Site directed mutagenesis of yeast hexokinase (Y-HKA) demonstrated the importance of Ser158 and Phe160, which are critical for glucose binding (Kraakman et al., 1999). These residues are altered in DHK-465 with alanine and tyrosine, respectively (Figure 2A). Whereas, DHK-453 sequence shows the conservation in these residues. In addition to the involvement of the amino acid residues of glucose binding domain, site directed mutagenesis of human HK-1, demonstrated that Asp657, Glu708 and Glu742 residues were involved in glucose binding and catalysis (Arora et al., 1991). Mutation in Glu708 of HK-1 resulted in 50-fold increase in the km for glucose. Asp657 and Glu708 of HK-1 are altered to Val and Asp respectively in DHK-465. Whereas, Glu742 is conserved at 286 position in DHK-465. All these residues are conserved in DHK-453 sequence (Table 1).
Out
of 13 residues important in the ATP binding domain, DHK-465 shows alteration
in 5 residues (Figure 2B). Arg539
of human HK-1 has been shown to be essential for catalysis as it stabilizes
the transition state product ADP-HK (Zeng et al., 1998). It is conserved in DHK465 at Arg83. Mutation of Gly534 affected
ATP binding of HK-1 (Zeng et al., 1998). This
residue is altered in DHK-465 with a methionine. Earlier studies from the
site directed mutagenesis of human brain HK-1 have shown that Asp532 in the
ATP binding domain interacts with Mg2+ ion and Thr680 stabilize the g–phosphoryl
group of ATP, and their interactions are important for the stabilization of
the transition state (Zeng et al.,
1996). Both Asp532 and Thr680 are replaced by glutamate and serine in DHK-465,
respectively, (Figure 2B and Table 1). Mutations in Gly862 and Gly679 in human
HK-1 has a significant influence on the binding affinity of ATP (Zeng et
al., 1998). These residues are conserved in DHK-465. Most of the ATP binding residues are well
conserved in DHK-453 (Figure 2B and Table 1) as well as other Drosophila hexokinases, Hex C (AF2347469 and AJ309864) and Hex
A (AAF46507, AAG23047) (Data not shown).
|
A. Putative Substrate binding sites of DHK-465 and DHK-453
were compared with Drosophila
HKs (HexA; HexC), mammalian
(Human HK-1) and yeast
hexokinases (Y-HKA; Y-HKB). The important residues are marked with ‘*’ DHK-465 138 LPLGIAFAFTLKKLALDVGILVSWTKEF H-HK-1 138 MPLGFTFSFPCQQTSLDAGILITWTKGF B. Core sequence of ATP
binding domain of other hexokinases was compared with putative DHK-465
and DHK-453 domains. Highly conserved amino acids in this domain are
marked with ‘*’ DHK-465 73 LALEMMPTNCRIMLV Y-HKA 83 LAIDLGGTNLRVVLV Figure 2. Comparison of putative substrate binding
domain and ATP binding domain of DHK with other hexokinases. |
|
Table1. The
comparison of amino acid residues important in substrate
§ Identical residues between HK-1
and DHK-465; * Identical residues |