
Weisman, N.Ya., I.D. Erokhina, and I.K. Zakharov. 2001. Concentration of net-extra-analis mutation in Drosophila melanogaster populations from nature in 2000-2001. Dros. Inf. Serv. 84: 117-120.
|
|
|
|||
|
|
||||
Concentration
of net-extra-analis mutation in Drosophila melanogaster populations
from nature in 2000-2001.
Weisman, N.Ya., I.D. Erokhina, and
I.K. Zakharov. Institute
of Cytology and Genetics of Siberian Branch of the Russian Academy of Sciences,
Novosibirsk, 630090, Russia; e-mail:
weisman@bionet.nsc.ru
Mutations in
the net gene (2-0.0) are known to disrupt the pattern of the wing
veining. Currently, about two dozen spontaneous and induced alleles of this
gene are known (Lindsley and Zimm, 1992; Biehs et al., 1998; Brentrup et al., 2000). As a rule, analogously to plexus mutations,
the net mutations lead to formation of a net of wing veins, as well
as to appearance of additional longitudinal fragments of veins in the wing
cells. Disruption of veining appears in all the wing cells, except
the first dorsal ones, which are characterized by expansion to a distal part.
The veins L4 and L5, beginning from the posterior cross-vein, are frequently
joined together.
In different
net alleles, the variation is observed in intensity of expression
of the mutant phenotype. In heterozygotes
with the normal allele, almost all known net-mutations
are recessive, except the net-41e4 allele.
In this case, in half of the individuals with the genotype net41e4
/ net+, the small portion of additional
veins could be registered, especially in the region L3 (Neel, 1942).
From the Belokurikha (Altai) population from nature in 2000, we have
isolated an allele of the net locus (net-B27) with unusual
properties. The phenotype of
this mutation is characterized by an additional fragment of vein in the third
dorsal wing cell, near by the middle of the longitudinal anal vein (Figure 1b). In the flies
homozygous for netB27, the mutation was incompletely penetrant
(about 80%). The expression of the mutation varied
relative to the length of an additional vein. With respect to the normal allele
of the net locus, the netB27 mutation is recessive.
The phenotype
of individuals heterozygous for the Altai allele of netB27 and the standard net allele from the collection of the Laboratory
of Genetics of Populations of the Institute of Cytology and Genetics SB RAS,
which produces multiple disruptions of veining in homozygotes (Figure 1a), is similar to the phenotype of the
homozygotes netB27. Unlike netB27
/ netB27, an additional vein fragment
in netB27 / net appears in all
the individuals and has a somewhat larger length than in mutant homozygotes. In netB27 / net, the additional fragment of the vein
is sometimes formed as a small branch deviating from the posterior cross-vein
in the third dorsal wing cell. Thus, the netB27 allele dominates over the standard mutant net allele.
All the individuals
with the phenotype netB27 / Df(2)net62, where Df(2)net62-chromosome 2 carries a deletion in the region 21А1-21В4-5 involving the net and giant
larvae genes (Korochkina, Golubovsky, 1978), have the additional fragments of veins or branches in the second and third
dorsal wing cells and in the distal part of marginal and sub-marginal wing
cells.
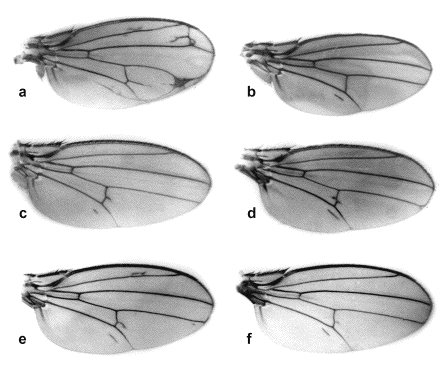
Figure 1. Variants of net-extra-analis phenotypes in homozygous and heterozygous with Df(2)net62 and net. а. net; b. net-extra-analisВ27; с. net-extra-analisU87; d net-extra-analisZ14; e. net-extra-analisZ14 / Df(2)net62; f. net-extra-analisZ14 / net.
The net
mutations with
the phenotype similar to the netB27 phenotype were found in all regions in geographically
remote populations studied by us in 2000 and 2001. Mutations were detected both in the flies
caught directly from nature and in the offspring of female inseminated in
the wild. For example, concentration of the net-chromosomes
in population from Pyuchas in 2001 has reached the level of 8%. The homozygotes
by different net alleles isolated from populations were
characterized by somewhat differing variants of mutant phenotypes. Except
the fragment of an additional vein in the third dorsal cell, we have observed
the branching or fragments of veins in the other cells in location of the
posterior cross-vein (Figure 1d). In heterozygotes with Df(2)net62
and the standard allele net, the mutations isolated from nature
were similar as netB27 (Figure 1e, f). According
to phenotypic penetration, the net mutation disrupting
the veining pattern near by the posterior cross-vein was termed by us as net-extra-analis.
|
By comparing the data of population from Pyuchas, Republic of Udmurtia, it is possible to note that in 2001, the concentration of the net-extra-analis mutation has increased from 2% to 8% (the sample number from populations is about 200 second chromosomes). Possibly, this genetic drift is related to the “founder” effect that appeared due to limited number of individuals in the studied population from nature, as well as due to strong seasonal fluctuations in the number of individuals.
Previously, in populational and genetical studies performed in the Laboratory of Genetics of Populations of IC&G SB RAS, the mutations with the similar phenotypic penetration were found in different years in various regions in geographically remote populations: in Ukraine population from Uman in 1987 (Figure 1c), in Altai population from Pospelikha in 1992, in Tadjikistan population from Dushanbe in 1990. The mutations isolated from nature were maintained in culture as homozygous strains in the collection of the Laboratory, but these data were not studied from the genetical view point.
As was mentioned in the well-known publication by N.P. Dubinin with co-workers (Dubinin et al., 1937), in all studied in 1933-1935 populations from Novorossyisk, Tashkent, Dagestan, and Crimea, the inherited aberrations in the chromosome 2 were detected. These aberrations were the type as net-extra-analis and denoted by authors as extra-analis and plexus-II. Their penetrance was expressed as formation of additional short fragments of veins in the region of the posterior cross-vein; however, they were not localized.
Based on the evidence given above, we may conclude that net-extra-analis mutations are constantly present in a noticeable concentration in the wild populations of Drosophila melanogaster from nature.
Acknowledgments: The authors are grateful to A.V. Ivannikov,
K.V. Gunbin, and Ya.Ya. Sinyansky for collecting the samples of Drosophila
melanogaster from populations in nature and to G.V. Orlova for translating
the manuscript into English. The
work is supported by the Russian Foundation for Basic Research, grant No. 99-04-49743.
References: Biehs, B., M.A. Sturtevant, and E. Bier 1998, Development 125: 4245; Brentrup, D., H. Lerch, H. Jackle, and M. Noll 2000,
Development
127: 4729; Dubinin, N.P.,
D.D. Romashov, M.A. Heptner, and Z.A. Demidova 1937, Biologicheskij J., 6: 311; Korochkina, L.S., and M.D. Golubovsky
1978, Dros. Inf. Serv. 53: 197;
Lindsley, D.L., and G.G. Zimm 1992,
The Genome of Drosophila
melanogaster. San Diego, CA, Academic Press Inc.; Neel, J., 1942,
Dros. Inf. Serv., 16: 49.